Uusi lehti lehdessä Elämä esittää varhaisen katsauksen virustekijöistä, jotka liittyvät apoptoosin käynnistämiseen ja toteuttamiseen tartunnan saaneissa soluissa.
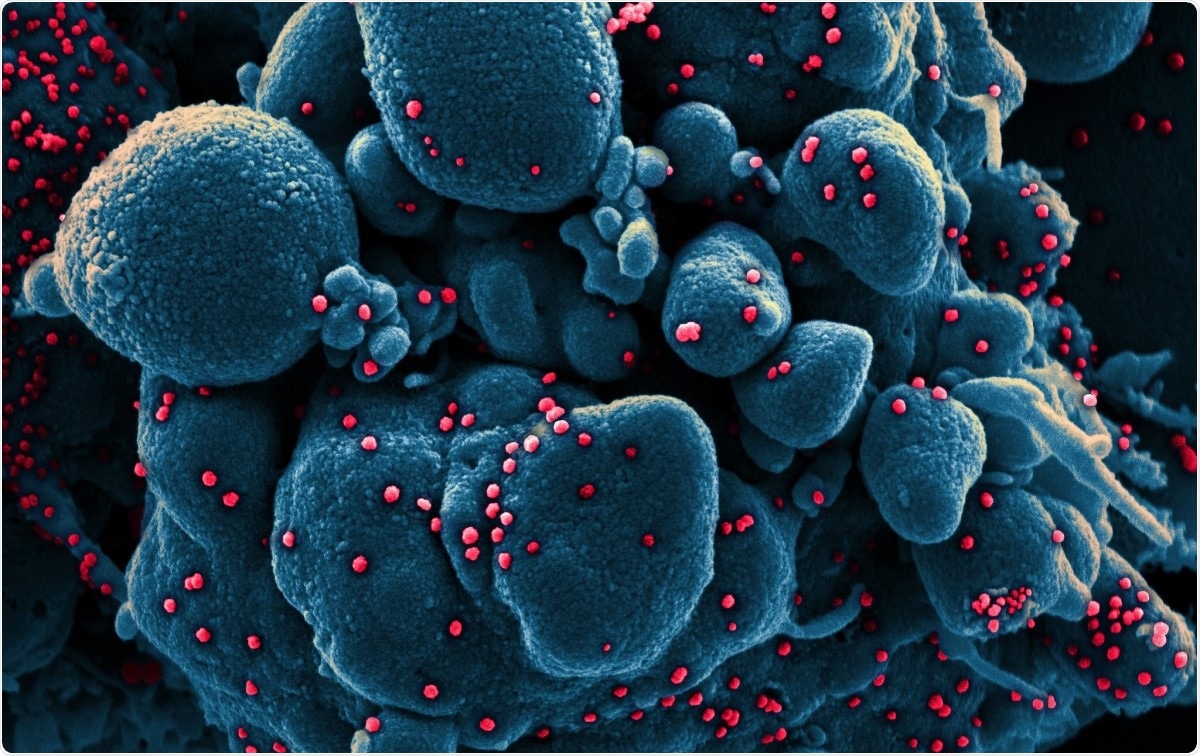 Värillinen pyyhkäisyelektronimikroskooppi SARS-COV-2-viruspartikkeleilla (punainen) tartunnan saaneesta apoptoottisesta solusta (sininen), eristetty potilasnäytteestä. Kuva otettu Fort Detrickin NIAID:n integroidussa tutkimuslaitoksessa (IRF), Maryland. Kuvaluotto:NIAID / Flickr
Värillinen pyyhkäisyelektronimikroskooppi SARS-COV-2-viruspartikkeleilla (punainen) tartunnan saaneesta apoptoottisesta solusta (sininen), eristetty potilasnäytteestä. Kuva otettu Fort Detrickin NIAID:n integroidussa tutkimuslaitoksessa (IRF), Maryland. Kuvaluotto:NIAID / Flickr Koronavirukset ovat suurimpia tähän mennessä tunnistettuja ribonukleiinihappoviruksia (RNA). Heidän genominsa on noin 30 kb pitkä, neljä rakenneproteiinia, ja useita ei-rakenteellisia ja lisäproteiineja. Ne luokitellaan alfaksi, beeta, gamma- ja delta -koronavirukset.
Useita eläinten ja ihmisten koronaviruksia on eristetty toistaiseksi ja ne aiheuttavat enimmäkseen hengitysteiden tai ruoansulatuskanavan infektioita. Kuten muutkin virukset, ne kaappaavat isäntäsolukoneiston replikoinnin kaikissa vaiheissa. Tähän sisältyy eri isäntägeenien ilmentymisen muuttaminen, mukaan lukien ne, jotka koodaavat isännän puolustusta viruksen replikaation helpottamiseksi.
Äskettäin, tutkimukset ovat osoittaneet, että infektion alkuvaiheessa, koronavirukset muuttavat solujen aineenvaihduntaa ja lisäävät solukuolemien määrää. Tämä tapahtuu ennen kuin adaptiivinen immuniteetti laukeaa, ja tämä voisi osoittaa sen roolin tuottavassa virusinfektiossa.
Kaksi tärkeintä tähän modulaatioon liittyvää prosessia ovat apoptoosi ja solusyklin muokkaaminen. Apoptoosi laukaisee useita signalointireittejä. Apoptoosi voi vähentää viruksen leviämistä, mutta myös päinvastainen vaikutus voi ilmetä - tarttuvien äskettäin tuotettujen viruspartikkeleiden vapautuminen lyyttisen infektion aikana.
Apoptoosin aktivoivat solupolut voivat laukaista suoraan virusinfektio itse tai epäsuorasti muiden signalointikaskadien kautta. Jälkimmäisen laukaisee viruksen läsnäolo ja rekrytoida immuunisoluja infektiokohtaan. Lopputapahtuma on apoptoosi, aiheuttaen merkittävää vahinkoa kudoksille lyyttisessä infektiossa.
Solun sisällä olevia viruskomponentteja kutsutaan patogeeniin liittyviksi molekyylikuvioiksi (PAMP) ja ne tunnistetaan kuvion tunnistusreseptoreista (PRR). Nämä reagoivat aktivoimalla sytokiinejä ja muita signalointimolekyylejä, jotka säätelevät interferonigeenin ilmentymistä.
Toinen puolustusmekanismi sisältää viruksen nukleiinihapon tunnistamisen isäntäproteiineilla, jotka liittyvät, kuten PRR:t, adapteriproteiineilla kahden keskeisen transkriptiotekijän aktivoimiseksi, nimittäin, NFĸB- ja IFN -sääntelytekijä 3 (IRF3). Nämä siirretään ytimeen ja lisäävät tyypin I interferonien (IFN) ilmentymistä.
Nämä sitoutuvat IFN-a/β-reseptoriin (IFNAR), vuorostaan, käynnistää signaalijärjestelmän, johon kuuluu Janus -kinaasi/signaalimuuntimet ja transkription aktivaattorit (JAK/STAT). Tuloksena on useiden IFN-stimuloitujen geenien (ISG) ylössäätely.
Nämä geenit inaktivoivat soluproteiinien synteesiin liittyvän prosessin ja aiheuttavat siten solukuoleman. Toinen ISG, joka on toimija tässä käännöksen pysäyttämisprosessissa, on yhdistelmä 2 ′, 5′-oligoadenylaattisyntetaasi (OAS) ja RNaasi L.
Apoptoosia säätelevät useat pro- ja anti-apoptoottiset proteiinit isäntäsolussa. Yksi tällainen proteiiniperhe on Bcl-2 (B-solulymfooma-2) -perhe, mukaan lukien tekijät, jotka aiheuttavat mitokondrioiden kalvon poraation ja mahdollisen kaspaasientsyymien aktivoitumisen, aloittaa apoptoosin. Toiset suojaavat apoptoosilta.
Toinen perhe on mitogeeniaktivoidut proteiinikinaasit (MAPK), jotka sisältävät solunulkoisen säädellyn kinaasin 1/2 (ERK1/2), ERK5, s38, ja c-Jun N-terminaaliset kinaasit (JNK). Ensimmäistä säätelevät kasvutekijät ja mitogeenit, kaksi viimeistä solustressin vuoksi. JNK-reitti voi johtaa pro-apoptoottisten geenien aktivoitumiseen tai voi olla vuorovaikutuksessa Bcl-2-geenien kanssa, tai voi aktivoida muita transkriptiotekijöitä, kuten 53 ja p73.
Apoptoosia on tunnistettu kaksi, ulkoinen ja sisäinen. Edellistä välittää kuoleman ligandien (FasL, TNF) niiden vuorovaikutuksesta vastaavien kuolemareseptoreidensa (DR) kanssa. Kaspaasi-8 on merkittävä apoptoosin indusoija eläinten koronaviruksissa, jotka vaikuttavat DR:ien kautta. Tämä entsyymi, aktivoituna, katkaisee tarjouksen, Bcl2 -proteiini, mikä johtaa mitokondrioiden sytokromi c:n vapautumiseen. Jälkimmäinen on avain apoptosomin muodostumiseen pilkkomalla ja aktivoimalla kaspaasi 9.
Luontainen reitti perustuu mitokondrioiden ulkokalvojen läpäisevyyteen ja nähdään pääasiassa eläinten koronaviruksilla. Bcl2-proteiinit, kuten Bax ja Bak, suosivat apoptoosia ja niiden on käynnistettävä tämä prosessi luontaisen tai mitokondrioiden välittämän reitin kautta, yhdessä kaspaasi-9:n kanssa.
Monet eläinten koronavirukset aktivoivat sekä ulkoisia että sisäisiä reittejä, joka esittää apoptoosiin liittyvän signalointireittien monimutkaisen verkon.
Virusgenomit koodaavat usein geenejä, jotka ovat homologisia isännän anti-apoptoottisille proteiineille. Tämä sallii niiden viivästyttää apoptoosia infektion alkuvaiheessa ja siten tuottaa riittävän määrän uusia virioneja. Muut virusproteiinit voivat estää joidenkin apoptoottisten geenien transkription tai aiheuttaa niiden proteiinituotteiden modifioinnin, tukahduttaakseen toimintansa.
Virusproteiinien aiheuttama apoptoosin indusointi voi sisältää korkki-riippuvan translaation lopettamisen isäntäsoluissa, indusoi apoptoosia, kuten polioviruksen kohdalla. Hepatiitti C -viruksen tapauksessa viruksen NS3-proteiini on vuorovaikutuksessa kaspaasi-8:n kanssa, laukaista solukuolema.
Nykyinen katsaus keskittyy tunnistamaan pro-apoptoottisia mekanismeja, jotka ovat aiheuttaneet eläinten koronavirukset. Esimerkiksi, Sian epidemian ripulin virus (PEDV) laukaisee muutoksia 14 geenin ilmentymisessä, jotka osallistuvat kaspaasista riippumattomaan apoptoosiin sen replikaation edistämiseksi, sekä tuottaa taudille ominaisia piirteitä.
Toinen tapa, jolla apoptoosi osallistuu viruksen replikaatioon, on aktivoimalla kaspaasi, jota tarvitaan viruksen nukleokapsidi (N) -proteiinin pilkkoutumiseen tämän prosessin aikana. Tämä koskee tarttuvaa gastroenteriittivirusta (TGEV), joka aktivoi MAPK -perheen, sekä FasL- ja mitokondrioiden aiheuttama apoptoosi. Tämä prosessi sisältää myös isäntämikroRNA:iden alareunan, jotka tukahduttavat mitokondrioiden apoptoottisen reitin.
Betacoronavirus, kuten hiiren hepatiittivirus (MHV), aiheuttaa apoptoosia. Tämä virus osoittaa kirjekuoriproteiinin (E) aiheuttamaa apoptoosia, uudelleen kaspaasin aktivoinnin kautta. Tässä tapauksessa, apoptoosi näyttää suojaavan virionituotantoa vastaan.
Sian hemagglutinoivan enkefalomyeliittiviruksen (PHEV) kanssa, solukuolemaa välittävät DR:t ja mitokondrioiden välittämät reitit, johon kuuluu kaspaasi-8 ja kaspaasi-9, aloittajina, ja kaspaasi-3, apoptoosin tekijänä.
Samoin, gamma- ja delta-koronavirukset indusoivat monimutkaisia solun signalointireittejä isäntäsolussa, mikä johtaa apoptoosin modulaatioon. Täten, monet virusproteiinit moduloivat isäntäsolureittejä edistääkseen viruksen replikaatiota ja taudin ominaisuuksia.
Se tosiasia, että apoptoosi tapahtuu useiden isäntien useiden koronavirusten aiheuttaman tartunnan jälkeen, johtaa siihen johtopäätökseen, että se on yleinen piirre ihmisten ja eläinten koronavirusten aiheuttamalle tartunnalle ja pyrkii edistämään leviämistä.
Aiemmin vakavan akuutin hengitystieoireyhtymän koronaviruksen (SARS-CoV) kanssa, 7a-proteiini muodostaa kompleksin Bcl-XL-proteiinin tai muiden selviytymistä edistävien tekijöiden kanssa, inaktivoimalla ne ja sallien siten apoptoosin etenemisen kaspaasi-3-aktivaation kautta. 3CL:n kaltainen proteaasi (3CLpro) aktivoi myös sekä kaspaasi-3:n että kaspaasi-9:n indusoimaan apoptoosia soluviljelmässä.
Sekä SARS-CoV- että tällä hetkellä kiertävillä SARS-CoV-2-viruksilla on ORF3a-proteiini, joka laukaisee solukuoleman kaspaasiaktiivisuuden kautta DR:ien läpi kulkevassa ulkoreitissä.
Samanlaista roolia ei ole vielä osoitettu virusproteiineilla eläinten koronavirusinfektioissa. Kuitenkin, PEDV:llä, piikkiproteiinin S1 -alayksikkö, ja ei-rakenteellinen proteiini 1 (NSP1), on ehdotettu solukuoleman käynnistäjäksi ja toteuttajaksi, vastaavasti, viimeaikaisten kokeilujen perusteella.
Lisää tutkimuksia voi auttaa ymmärtämään, miten nämä reitit toimivat muissa koronaviruksissa. Tämänhetkinen tietämys voi osoittaa, kuinka hyödyllistä on kohdistaa tämä reitti koronavirusinfektioiden hallintaan.
 Eräs suolistobakteeri voi lisätä suolistosyövän riskiä
Eräs suolistobakteeri voi lisätä suolistosyövän riskiä
 Miten isäntätekijät, kuten keuhkojen mikrobiomi, auttavat SARS -CoV -2 -infektiossa?
Miten isäntätekijät, kuten keuhkojen mikrobiomi, auttavat SARS -CoV -2 -infektiossa?
 Yksityiskohtainen kartta ihmisen kielen mikrobiomista
Yksityiskohtainen kartta ihmisen kielen mikrobiomista
 Metformiini voi auttaa vuotavaa suolistoa
Metformiini voi auttaa vuotavaa suolistoa
 Punkit kantavat nyt useita sairauksia,
Punkit kantavat nyt useita sairauksia,
 Valkosolut ja niiden rooli aivoissa
Valkosolut ja niiden rooli aivoissa
 Keuhkomikrobit voivat auttaa ennustamaan vakavasti sairaiden tuloksia
Uusi tutkimus julkaistiin American Journal of Respiratory and Critical Care Medicine osoittaa, että keuhkoissa elävien organismien tyypin ja lukumäärän muuttumismuodon kirjaaminen voisi heijastaa va
Keuhkomikrobit voivat auttaa ennustamaan vakavasti sairaiden tuloksia
Uusi tutkimus julkaistiin American Journal of Respiratory and Critical Care Medicine osoittaa, että keuhkoissa elävien organismien tyypin ja lukumäärän muuttumismuodon kirjaaminen voisi heijastaa va
 Muuttoliike vaikuttaa suoliston mikrobistoon, mikä puolestaan vaikuttaa tutkijoiden terveyteen
Ihmisen suolistossa on biljoonia hyödyllisiä ja ystävällisiä bakteereja, jotka muodostavat mikrobiomin. Näiden mikrobien havaitaan vaikuttavan suuresti ihmisten olosuhteisiin, mukaan lukien yleinen te
Muuttoliike vaikuttaa suoliston mikrobistoon, mikä puolestaan vaikuttaa tutkijoiden terveyteen
Ihmisen suolistossa on biljoonia hyödyllisiä ja ystävällisiä bakteereja, jotka muodostavat mikrobiomin. Näiden mikrobien havaitaan vaikuttavan suuresti ihmisten olosuhteisiin, mukaan lukien yleinen te
 OTC-antasidien hapon neutralointikyvyn määrittäminen
USP kuvaa vasta-titrauksen happoa neutraloivan kapasiteetin (ANC) määrittämiseksi. Menetelmä on erittäin nopea, helppo, ja luotettava. Metrohm-sovellushuomautus AN T-192 sisältää ANC:n tehokkaan määr
OTC-antasidien hapon neutralointikyvyn määrittäminen
USP kuvaa vasta-titrauksen happoa neutraloivan kapasiteetin (ANC) määrittämiseksi. Menetelmä on erittäin nopea, helppo, ja luotettava. Metrohm-sovellushuomautus AN T-192 sisältää ANC:n tehokkaan määr