Et nytt papir i journalen Liv presenterer en tidlig gjennomgang av de virale faktorene som er involvert i å starte og utføre apoptose i infiserte celler.
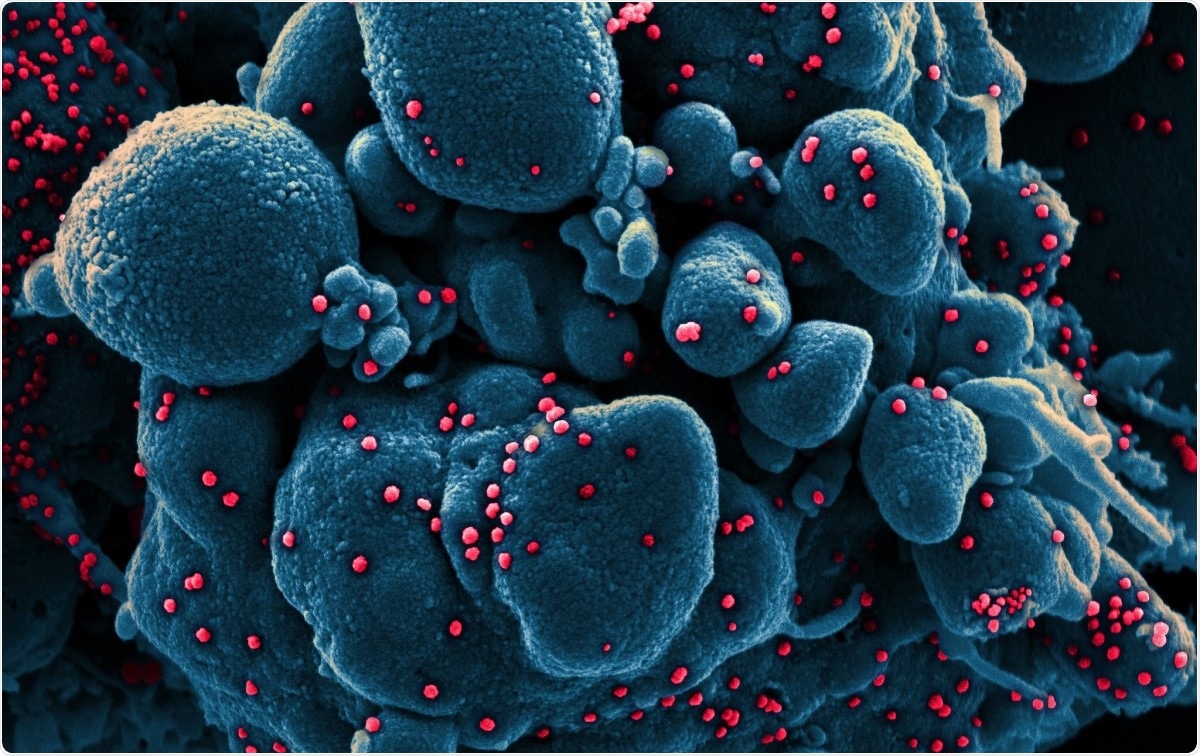 Farget skanningelektronmikrograf av en apoptotisk celle (blå) infisert med SARS-COV-2-viruspartikler (rød), isolert fra en pasientprøve. Bilde tatt på NIAID Integrated Research Facility (IRF) i Fort Detrick, Maryland. Bildekreditt:NIAID / Flickr
Farget skanningelektronmikrograf av en apoptotisk celle (blå) infisert med SARS-COV-2-viruspartikler (rød), isolert fra en pasientprøve. Bilde tatt på NIAID Integrated Research Facility (IRF) i Fort Detrick, Maryland. Bildekreditt:NIAID / Flickr Koronavirus er blant de største ribonukleinsyre (RNA) virusene som er identifisert så langt. Genomet deres er omtrent 30 kb langt, med fire strukturelle proteiner, og flere ikke-strukturelle og tilbehørsproteiner. De er klassifisert i alfa, beta, gamma- og delta -koronavirus.
Flere koronavirus fra dyr og mennesker har blitt isolert så langt og forårsaker hovedsakelig luftveis- eller gastrointestinale infeksjoner. Som andre virus, de kaprer vertscelle -maskineriet i alle stadier av replikasjonen. Dette innebærer å modifisere uttrykket for ulike vertsgener, inkludert de som koder vertsforsvar for å lette virusreplikasjon.
Nylig, studier har vist at tidlig i infeksjonen, koronavirus endrer cellemetabolismen og øker celledødsfrekvensen. Dette skjer før adaptiv immunitet utløses, og dette kan indikere dens rolle i produktiv virusinfeksjon.
To av de viktigste prosessene som er involvert i denne modulasjonen er apoptose og cellesyklusmodifikasjon. Apoptose utløses av en rekke signalveier. Apoptose kan redusere virusutbredelse, men den motsatte effekten kan også forekomme - frigjøring av de smittsomme nyproduserte viruspartiklene under lytisk infeksjon.
Celleveiene som aktiverer apoptose kan utløses direkte av selve virusinfeksjonen eller indirekte via andre signalkaskader. Sistnevnte motvirkes av tilstedeværelsen av viruset og rekrutterer immunceller til infeksjonsstedet. Sluttbegivenheten er apoptose, forårsaker betydelig skade på vevet ved lytisk infeksjon.
De virale komponentene i cellen kalles patogenassosierte molekylære mønstre (PAMP) og gjenkjennes av mønstergjenkjenningsreseptorer (PRR). Disse reagerer ved å aktivere cytokiner og andre signalmolekyler som oppregulerer uttrykk for interferon -gen.
En annen forsvarsmekanisme innebærer gjenkjenning av viral nukleinsyre av vertsproteiner som forbinder, som PRR, med adapterproteiner for å aktivere to viktige transkripsjonsfaktorer, nemlig, NFĸB og IFN regulatorisk faktor 3 (IRF3). Disse translokeres til kjernen og øker ekspresjonen av type I interferoner (IFN).
Disse binder seg til IFN-α/β reseptor (IFNAR), i sin tur, sette i gang et system med signaler som involverer Janus kinase/signaltransdusere og aktivatorer for transkripsjon (JAK/STAT). Resultatet er oppregulering av flere IFN-stimulerte gener (ISG).
Disse genene inaktiverer prosessen som er involvert i syntesen av celleproteiner og induserer dermed celledød. En annen ISG som er en spiller i denne prosessen med å stenge oversettelse er kombinasjonen av 2 ′, 5′-oligoadenylatsyntetase (OAS) og RNase L.
Apoptose reguleres av flere pro- og anti-apoptotiske proteiner i vertscellen. En slik proteinfamilie er familien Bcl-2 (B-cellelymfom-2), inkludert faktorer som forårsaker porering av mitokondriemembranen og eventuell aktivering av caspasasezymer, starter apoptose. Andre beskytter mot apoptose.
En annen familie er mitogenaktiverte proteinkinaser (MAPK), som inkluderer ekstracellulær regulert kinase 1/2 (ERK1/2), ERK5, p38, og c-Jun N-terminale kinaser (JNK). Den første er oppregulert av vekstfaktorer og mitogener, de to siste av cellestress. JNK-banen kan føre til aktivering av pro-apoptotiske gener eller kan samhandle med Bcl-2-gener for å gjøre det, eller kan aktivere andre transkripsjonsfaktorer som 53 og p73.
To veier for apoptose er identifisert, det ytre og det indre. Førstnevnte formidles av aktivering av dødsligander (FasL, TNF) om deres interaksjon med sine respektive dødsreseptorer (DR). Caspase-8 er en viktig induktor for apoptose i dyrekoronavirus som virker via DRs. Dette enzymet, når den er aktivert, kløver bud, et Bcl2 -protein, som fører til frigjøring av mitokondrielt cytokrom c. Sistnevnte er nøkkelen til dannelsen av apoptosomet med spaltning og aktivering av caspase 9.
Den iboende banen er basert på permeabilisering av mitokondriale ytre membraner og er hovedsakelig sett med koronavirus fra dyr. Bcl2-proteiner som Bax og Bak favoriserer apoptose og er påkrevd for å utløse denne prosessen via den indre eller mitokondriemedierte banen, sammen med caspase-9.
Mange koronavirus fra dyr forårsaker aktivering av både ytre og indre veier, viser det intrikate nettverket av signalveier involvert i apoptose.
Virale genomer koder ofte for gener som er homologe for å være vert for anti-apoptotiske proteiner. Dette lar dem forsinke apoptose i de tidlige stadiene av infeksjonen og dermed generere et tilstrekkelig utbytte av nye virioner. Andre virale proteiner kan forhindre transkripsjon av noen apoptotiske gener eller forårsake endring av proteinproduktene deres, for å undertrykke deres aktivitet.
Induksjon av apoptose av virale proteiner kan innebære avslutning av cap-avhengig translasjon i vertsceller, indusere apoptose, som sett med poliovirus. Når det gjelder hepatitt C -virus, det virale NS3-proteinet interagerer med caspase-8, å utløse celledød.
Den nåværende anmeldelsen er fokusert på å identifisere pro-apoptotiske mekanismer utløst av dyrekoronavirus. For eksempel, Porcine Epidemic Diarrhea Virus (PEDV) utløser endringer i uttrykket for 14 gener som er involvert i caspase-uavhengig apoptose for å fremme replikasjonen, i tillegg til å produsere de karakteristiske trekk ved sykdom.
En annen måte apoptose er involvert i virusreplikasjon på, er ved å aktivere caspase, som er nødvendig for spaltning av det virale nukleokapsid (N) -proteinet under denne prosessen. Dette er tilfellet med det overførbare gastroenterittviruset (TGEV), som aktiverer MAPK -familien, samt FasL- og mitokondrier-indusert apoptose. Denne prosessen inkluderer også nedregulering av vertsmicroRNA som undertrykker den mitokondrielle apoptotiske banen.
Betacoronavirus, slik som murin hepatittvirus (MHV), induserer apoptose. Dette viruset viser konvoluttprotein (E) -indusert apoptose, igjen via caspase -aktivering. I dette tilfellet, apoptose ser ut til å være beskyttende mot virionproduksjon.
Med svinehemagglutinerende encefalomyelittvirus (PHEV), celledød medieres av DRs og de mitokondriemedierte veiene, som involverer caspase-8 og caspase-9, som initiativtakere, og caspase-3, som en effektor av apoptose.
På samme måte, gamma- og delta-koronaviruset viser kompleks induksjon av cellulære signalveier i vertscellen, som fører til modulering av apoptose. Og dermed, mange virale proteiner modulerer vertscelleveier for å fremme viral replikasjon og sykdomsfunksjoner.
Det faktum at apoptose oppstår etter infeksjon av flere verter med en rekke koronavirus, fører til konklusjonen at det er et vanlig trekk ved infeksjon av mennesker og dyr og har som mål å favorisere forplantning.
Med tidligere alvorlig akutt respiratorisk syndrom coronavirus (SARS-CoV), 7a-proteinet danner et kompleks med Bcl-XL-protein eller andre pro-overlevelsesfaktorer, inaktivere dem og dermed tillate apoptose å fortsette via caspase-3-aktivering. Den 3CL-lignende protease (3CLpro) aktiverer også både caspase-3 og caspase-9 for å indusere apoptose i cellekultur.
Både SARS-CoV og de for tiden sirkulerende SARS-CoV-2-virusene har ORF3a-proteinet som utløser celledød via caspaseaktivering i den ekstrinsiske banen gjennom DRs.
En lignende rolle er ennå ikke vist for virale proteiner ved infeksjoner av koronavirus fra dyr. Derimot, med PEDV, S1 -underenheten til piggproteinet, og det ikke-strukturelle protein 1 (NSP1), har blitt foreslått å være initiativtaker og effektor av celledød, henholdsvis basert på nylige forsøk.
Flere studier kan hjelpe deg med å forstå hvordan disse veiene fungerer i andre koronavirus. Den nåværende kunnskapstilstanden kan indikere nytten av å målrette denne veien for å håndtere koronavirusinfeksjoner.
 Vanlig genetisk variant forklarer hvorfor immunterapi ofte mislykkes ved Crohns sykdom
Vanlig genetisk variant forklarer hvorfor immunterapi ofte mislykkes ved Crohns sykdom
 Vekstproblemer hos premature spedbarn assosiert med endrede tarmbakterier
Vekstproblemer hos premature spedbarn assosiert med endrede tarmbakterier
 Hurtigmat kan være den viktigste synderen i tenåringsdepresjon
Hurtigmat kan være den viktigste synderen i tenåringsdepresjon
 Mat påvirker selektivt tarmmikrober, finner studier
Mat påvirker selektivt tarmmikrober, finner studier
 Probiotika kan bidra til å dempe underernæring de neste to tiårene,
Probiotika kan bidra til å dempe underernæring de neste to tiårene,
 Munnhygiene og alvorlighetsgraden av COVID-19-forbindelsen
Munnhygiene og alvorlighetsgraden av COVID-19-forbindelsen
 Enkelte bakteriearter kan øke HIV -risikoen hos kvinner,
finner ny studie En nylig studie publisert i The Lancet Infectious Diseases, beskriver syv arter av vaginale bakterier som kan øke risikoen for HIV -infeksjon betydelig hos kvinner. K
Enkelte bakteriearter kan øke HIV -risikoen hos kvinner,
finner ny studie En nylig studie publisert i The Lancet Infectious Diseases, beskriver syv arter av vaginale bakterier som kan øke risikoen for HIV -infeksjon betydelig hos kvinner. K
 Vibrasjoner i hele kroppen bidrar til å redusere betennelse,
takket være tarmmikrobiomet Vibrasjoner i hele kroppen ser ut til å forbedre mange symptomer på type II diabetes mellitus, der glukose og destruktiv betennelse skyter opp. Prosedyren hjelper kroppen m
Vibrasjoner i hele kroppen bidrar til å redusere betennelse,
takket være tarmmikrobiomet Vibrasjoner i hele kroppen ser ut til å forbedre mange symptomer på type II diabetes mellitus, der glukose og destruktiv betennelse skyter opp. Prosedyren hjelper kroppen m
 Sædmikrobiom avslørt med RNA -sekvensering
En ny studie rapporterer den første noensinne detaljerte beskrivelsen av det menneskelige sædmikrobiomet, ved bruk av nyere RNA -sekvenseringsteknikker som er i stand til å skille mellom sæd -RNA og b
Sædmikrobiom avslørt med RNA -sekvensering
En ny studie rapporterer den første noensinne detaljerte beskrivelsen av det menneskelige sædmikrobiomet, ved bruk av nyere RNA -sekvenseringsteknikker som er i stand til å skille mellom sæd -RNA og b