absztrakt katalógusa
Háttér katalógusa
A gyomorrák továbbra is az egyik leghalálosabb rák a világon, és ezért azonosítása új gyógyszerek célba ez a fajta rák tehát nagy jelentősége. A tanulmány célja az volt, hogy azonosítsa és ellenőrizze terápiás szer, amely javítja a kimenetelt gyomorrákos betegek a jövőben. Katalógusa
Módszertan /fő eredményei katalógusa
A microarray technológia, hoztunk létre egy olyan gént expressziós profilja humán gyomorrák-specifikus gének a humán gyomorrák szövetmintákban. Használtuk ezt a profilt a Broad Intézet kapcsolati térkép elemzés azonosítani jelölt terápiás vegyületeket gyomorrák. Azt találtuk, hogy a hiszton dezacetiláz inhibitor Vorinostat, mint az ólom vegyület, és így egy potenciális terápiás gyógyszer gyomorrák. Vorinostat indukált apoptózist és autofágia gyomor rákos sejtvonalak. Farmakológiai és genetikai gátlása autofágia azonban növekedett a terápiás hatékonyságát vorinostat, jelezve, hogy a kombináció Vorinostat együtt autofágia inhibitorok terápiásán előnyösebb. Ezen kívül, a génexpressziós analízise az gyomorrák azonosított gyűjteménye gének ( ITGB5, TYMS, MYB, APOC1, CBX5, PLA2G2A, Következtetések /Jelentés katalógusa Kimutattuk, hogy elemzése génexpressziós aláírás képviselhet egy feltörekvő megközelítés felfedezni terápiás szerek gyomorrák, mint például a Vorinostat. A megfigyelés megváltozott génexpresszió után vorinostat kezelés biztosítja a nyom, hogy azonosítsa a molekuláris mechanizmusa vorinostat és azok a betegek számára hasznosak lehetnek az vorinostat kezelést. Katalógusa Citation: CLAERHOUT S, Lim JY, Choi W, Park YY, Kim K, Kim SB, et al. (2011) Gene Expression Signature analízis megállapítja Vorinostat jelöltként Therapy gyomorrák. PLoS ONE 6 (9): e24662. doi: 10,1371 /journal.pone.0024662 katalógusa Vágó: David L. McCormick, IIT Kutató Intézet, az Amerikai Egyesült Államok katalógusa Beérkezett: március 29, 2011; Elfogadva: augusztus 16, 2011; Megjelent: szeptember 9, 2011 katalógusa Copyright: © 2011 CLAERHOUT et al. Ez egy nyílt hozzáférésű cikk feltételei szerint terjeszthető a Creative Commons Nevezd meg! Licenc, amely engedélyezi a korlátlan használatát, a forgalmazás és a reprodukció bármilyen adathordozón, feltéve, hogy az eredeti szerző és a forrás jóváírásra. Katalógusa Forrás: finanszírozása SC mint Odyssey Fellow támogatta az Odyssey Program és a Theodore N. törvény Alapítvány tudományos teljesítmény, a University of Texas MD Anderson Cancer Center. Ez a munka is támogatta alapok a 2009 Internal Medicine Academic Research Fund és a Basic Science Research Program a Nemzeti Kutatási Alapítvány Koreai Minisztérium támogatja az Oktatási, Tudományos és Technológiai (No. 2010-0024248). A finanszírozók nem volt szerepe a tanulmány tervezés, adatgyűjtés és elemzés, döntés, hogy közzéteszi, vagy a készítmény a kézirat. Katalógusa Érdekütközés: A szerzők kijelentették, hogy nem ellentétes érdekek léteznek. Katalógusa Bevezető a gyomorrák a negyedik leggyakoribb daganat a második vezető daganatos halálok a világon [1], a teljes túlélés körülbelül 10 hónap [2] - [4]. A kezelés a gyomorrák lehetnek a kemoterápia, műtét és sugárkezelés. Sajnos, a jelenlegi kemoterápiás kezeléseket az előrehaladott gyomorrák bizonyítani kiábrándító eredményei [2] - [4]. Valóban, teljes remisszió ritkák, vagy csak tart nagyon röviden. Katalógusa Számos célzott szerek biztosítanak túlélési előnyt más ráktípus nyomozás alatt a gyomorrák. Míg néhány korai klinikai vizsgálatokban a vaszkuláris endoteliális növekedési faktor receptor (VEGFR), és epiteliális növekedési faktor receptor (EGFR) -1-inhibitorok, mint például a cetuximab és bevacizumab, kimutatták, némileg aktivitás, ott ritkán egy tényleges túlélési előny a beteg számára [5] , [6]. Ennek egyik oka az lehet, hogy ezek a vizsgálatok nem betegek kiválasztására megléte szerint a biomarkerek. A közelmúltban, a Trastuzumab gyomorrák (ToGA) vizsgálatban megállapította, hogy a túl trastuzumab kemoterápiával vezetett statisztikailag szignifikáns javulást progressziómentes túlélés (PFS) és a teljes túlélés (OS) a mintegy 20% -ánál a disszeminált gyomor- és gastrooesophagealis (GE) csomópont tumorok overexpresszáló HER2 [7]. Ez hangsúlyozza annak szükségességét, hogy a célzott biológiai terápia és a keresést a biomarkerek betegek kiválasztására klinikai vizsgálatok, amelyek részesülhetnek a túlélést. Annak ellenére, hogy néhány bizonyíték a potenciális célpontok, beleértve a HER2 [8], [9] a hatásosságát ezeknek a biológiailag célzott terápiák nem ismert és hiányzik a szabványos célzott terápia gyomorrák. Miatt a biológiai heterogenitása gyomor rák, nem valószínű, hogy van egy "mágikus lövedék" gyógyítani. Molekuláris markerek lesz így a jövőben fontos megjósolni betegek eredményeit és a szabás kezelések, az egyedi biológia. Katalógusa A keresési biomarkerek génexpressziós aláírás elemzés használják a különböző alkalmazások, mint például megvilágítását lehetővé tevő mechanizmusai biológiai útvonalak [10], osztályozás altípusok a betegség [11], előre rák prognózis [12] és a profilozó génexpresszió adott válaszként a kábítószerek [13], [14]. A génexpressziós aláírás elemzés történhet a Broad Intézet kapcsolati térkép (http://www.broadinstitute.org/cmap). A Connectivity térkép célja, hogy létrehoz egy térképet, amely összekapcsolja génexpressziós járó betegség megfelelő minták által termelt gyógyszerjelöltek és génmanipulációs [15], [16]. Ez a rendszer a megközelítés lehetővé teszi szkrínelendő vegyületek ellen genomot betegség aláírások, nem pedig egy előre kiválasztott sor célgének. Kábítószer párosítva betegségek kifinomult mintaillesztéses módszerek magas szintű felbontást és specificitás. Annak ellenére, hogy hagy sok a nyitott kérdés, a kapcsolati térkép azt mutatja, hogy genetikai aláírás elemzés felhasználható felismerni gyógyszerek közös hatásmechanizmusát, felfedezni ismeretlen hatásmechanizmus és a potenciális új terápiás [15], [16]. Katalógusa a tanulmány célja az volt, hogy azonosítsuk a potenciális új terápiás kezelésére gyomorrák. Ehhez először elemezték a genom aláírása emberi gyomorrák. A kapott gyomorrák gén aláírás használjuk ezután in silico elemzése microarray adatok a Connectivity Map elemzés során a microarray adatok 65 gyomorrákos betegek, beleértve a 65 rákok és 19 normál gyomor szövetek, amelyeket kapott a mi korábbi munkája, Yonsei adatok [17]. Daganatmintákat gyűjtöttünk gyomorrákos betegek gastrectomián elsődleges kezelés. A szövetmintákat vizsgáltuk patológusok idején gyűjtés és tárolni -80 ° C-on a szövet bank megkezdéséig a kísérletet. Teljes RNS-t extraháljuk a frissen fagyasztott szövetek segítségével Mirvana RNS izolálás jelölő készlet (Ambion, Inc.). Elsődleges microarray adatok rendelkezésre állnak NCBI Gene Expression Omnibus nyilvános adatbázisban (microarray platform, GPL6884; microarray adatok, GSE 13861). Egy másik gén expressziós profiljának kapunk 69 gyomor szövetminták, köztük 38 a rák és a 31 nem rákos stroma, a Stanford Microarray Database (http://smd.stanford.edu, GSE13911), Stanford adatokat. A gyomorrák-specifikus géneket által kiválasztott BRB-ArrayTools verzió 3.6.1 (biometrikus Research Branch, National Cancer Institute, Bethesda, MD). Osztály összehasonlítás két mintás t-teszttel (szignifikancia < 0,001, 10.000 véletlen permutáció) azonosított gyomorrák specifikus géneket és a gének, melynek átlagos expressziós intenzitásokat megváltoztatták legalább két-szerese jelent normális szövet génexpressziót választottak ki. Kapcsolatok Map elemzés katalógusa a potenciális célzó gyógyszerek gyomorrák, a gén listák top 500-ig a szabályozott és a top 500 le gének a gyomorrák-specifikus géneket használják (táblázat S1). A Connectivity Térkép elemzést végeztünk a webes felületről (http://www.broadinstitute.org/cmap) alkalmazásával verzió, épít 02, amely több mint 7000 expressziós profilok képviselő hatásai 1309 vegyületek számos tenyésztett humán sejtek [15], [16]. A Connectivity térkép azt mutatja, funkcionális kapcsolatokat gyógyszerek közötti, a gének és a betegség. Gyógyszerek, amelyek betegséget okozhatnak utánzó gén aláírás segíthet azonosítani a pályákat, amik jelentenek potenciális terápiás célpontokat, amelyeket a betegség. Fordítva, gyógyszerek, amelyek indukálják a "fordított" aláírás, azaz változások a génexpresszió ellentétes irányban, hogy megfigyelhető a betegség stádiuma, jelenthet új terápiás szerek. Ágensjelöltek ellen egy adott betegség lehet ismerni alkalmazásával a betegség-specifikus génexpressziós profil a Kapcsolódás Map elemzés. Azért választottuk gyógyszerjelöltek érvényesítés in vitro katalógusa alapján a kapcsolat pontszám, korreláció, és a P-értéket. Katalógusa Vegyszerek és sejttenyészetben katalógusa Vorinostat vegyületet a Merck és elkészített törzsoldatként dimetil-szulfoxidban (DMSO). Klorokin, bafilomycin A1 (Sigma) feloldunk illetve a víz és a DMSO-val. Emberi gyomor rákos sejtvonalak AGS, NCI-N87, és KATO-III kaptuk az American Type Culture Collection, és tartottuk fenn aszerint, hogy azok ajánlásait. A sejteket RPMI 1640, kiegészítve 10% magzati borjúszérummal, 100 U penicillin és 100 ng /ml sztreptomicinnel, 37 ° C-on, 5% CO 2. a 3- (4,5-dimetil-tiazol-2-il) -2,5-difenil-tetrazólium-bromid (MTT) vizsgálat, MTT-t PBS-ben oldott 5 mg /ml volt. Körülbelül 5 × 10 3 sejteket oltottunk 96 lyukú lemezekre és éjszakán át hagytuk letapadni. A tápközeget ezután friss tápközeggel helyettesítettük, amely a megjelölt koncentrációjú Vorinostat vagy DMSO-ban. 72 óra elteltével 20 pl MTT-oldatot adtunk hozzá, és a lemezeket inkubáljuk 37 ° C hőmérsékleten 4 órán át. Az inkubálás után 100 ul DMSO-t adtunk, hogy feloldjuk a formazán, és abszorbancia leolvastuk 570 nm-en spektrofotometriás mikrotiterlemez-leolvasó (Vmax kinetikus mikrotiterlemez-leolvasó, Molecular Devices). A kísérleteket három párhuzamossal végzünk. kristály ibolya festést, a sejteket kezeltük 12 órán, a tápközeget eltávolítottuk, és a sejteket PBS-sel mostuk, majd 30 percig inkubáltuk 0,5% kristály ibolya (In 20% metanol és 80% kétszeresen desztillált víz). A sejteket ezután háromszor mossuk PBS-sel. A fennmaradó kristályibolya extraháltuk ecetsavval 5 percig, és abszorbancia mértük 595 nm-en spektrofotometriás mikrolemez leolvasóval. Annak meghatározására, a sejtek életképességét, a sejteket inkubáltuk Vorinostat 72 órán át. A letapadt sejteket ezután levált tenyésztő lemezek tripszinezéssel és kombinált úszó sejtek, centrifugáljuk és szuszpendáljuk 500 ul propidium-jodid (PI) -exclusive oldat (nem sejtmembrán áthatoló) 15 percen át 4 ° C-on. Megfestett sejteket követtük áramlási citometriával (Beckman Coulter Cytomics FC 500). sejtciklus analízis, sejteket inkubáltunk Vorinostat 24 órán, összegyűjtjük, és szuszpendáljuk 500 ul hipotóniás oldatban (0,1% nátrium-citrát, 0,1% Triton X-100, 100 ng /ml RN-áz és 50 ug /ml PI) 15 percen át 4 ° C-on. PI-festett sejteket nyomon áramlási citometriával. Sejtciklus elemeztük Multicycle AV szoftver. Katalógusa RNS izolálás és microarray kísérletek RNS izolálás és microarray kísérletet végeztünk protokoll szerint a leírtak szerint [17]. Teljes RNS-t extraháltunk gyomor rákos sejtvonalak, vagy anélkül Vorinostat kezelés alkalmazásával Mirvana RNS izolációs kit (Ambion, TX, USA). A integritását a nagy RNS-frakció segítségével határozzuk meg egy Experion Bioanalyzer (Bio-Rad, CA, USA), mint a helyettesítő mRNS minőség-ellenőrzés. Teljes RNS-t jelzett és hibridizáltuk humán HT12 V.3 expressziós BeadChips szerint a gyártó protokollja szerint (Illumina, CA, USA). Miután a BeadChips beszkennelt egy Illumina BeadArray Reader, a microarray adatok normalizálódtak a kvantilis normalizációs módszer a lineáris modellek microarray adatok csomagot az R nyelvi környezetben [18]. Az expressziós szintjét az egyes gének átalakult log 2 bázis előtt további elemzést. Klaszteranalízis végeztük Cluster és Treeview [19]. A sejteket lekapartuk a közepes és lecentrifugáljuk, és a fehérjéket izoláltunk lízis pufferben (50 mM HEPES, 150 mM nátrium-klorid, 1 mM EGTA-t és 10 mM nátrium-pirofoszfát (pH 7,4)), amely 100 mM NaF, 10% glicerin, 1,5 mM MgCI 2, 1% Triton X-100 és a proteáz inhibitor (Roche). Az extraktumokat jégen inkubáltuk 20 percig, és lecentrifugáltuk 20800 g-vel 20 percig. A fehérje koncentrációját alkalmazásával határoztuk meg BCA fehérje vizsgálati reagenssel (Pierce). Egyenlő mennyiségű fehérje minden mintából elkülönítettük végzett elektroforézissel SDS-PAGE és Hybond-C Super membránra (Amersham Pharmacia Biotech). A membránokat blokkoltuk 1 órán át szobahőmérsékleten Tris-pufferelt sóoldatban, amely 0,1% Tween-20 és 5% zsírmentes szárított tej. A membránokat egy éjszakán át inkubáltuk 4 ° C hőmérsékleten a primer antitesttel hígított 5% zsírmentes tejpor vagy 5% BSA-val 1 × Tris-pufferolt sóoldat plusz 0,1% Tween-20. Antitestek LC3 (Novus Biologicals), az aktív kaszpáz-3 (Epitomics) és P62 (BD Biosciences) használtunk. Antitestek az alfa-tubulin és Beclin-1 volt, a Cell Signaling Technology; antitest p-aktin volt a Sigma. A membránokat ezután mostuk, és 1 órán át inkubáltuk szobahőmérsékleten peroxidázzal konjugált másodlagos antitestet (Cell Signaling Technology). Protein tettük láthatóvá alkalmazásával fokozott kemilumineszcencia által leírtak szerint a gyártó (GE Healthcare). Az siRNS célszekvencia számára Beclin-1, nontargeting siRNS (Risc Free) és Dharmafect 1 voltak vásárolt Dharmacon. Sejteket szélesztettünk 10 cm-es ételek és transzfektált siRNS 24 órával később határoztuk meg a gyártó utasításai szerint. A következő napon a sejteket tripszinnel kezeltük, és beoltjuk a 6 cm-es vagy 96-lyukú lemezeken, így ugyanazt a transzfekció hatékonyságát. Protein expressziós szinteket határoztuk meg Western-blot-analízis. A mintákat fixáltuk tartalmazó oldattal 3% gluteraldehyde plusz 2% paraformaldehidet tartalmazó 0,1 M kakodilát pufferrel, pH = 7,3, 1 óra. Fixálás után, a mintákat mossuk, és az oldathoz 0,1% Millipore-szűrt kakodilát pufferolt csersav, utófixáltuk 1% -os pufferolt ozmium-tetroxidot, 30 percig, és megfestettük en bloc 1% Millipore-szűrt uranil-acetáttal extraháljuk. A mintákat dehidráltuk növekvő koncentrációjú etanolt, beszivárgott, és a beágyazott LX-112 tápközegben. A mintákat polimerizálunk egy 70 ° C-szárítószekrényben 2 napig. Ultravékony metszeteket egy Leica Ultracut mikrotom (Leica, Deerfield, IL), megfestettük uranil-acetáttal és ólom-citráttal egy Leica EM Stainer, és vizsgálandó JEM 1010 transzmissziós elektronmikroszkóppal (JEOL USA, Inc., Peabody, MA ) gyorsító feszültség 80 kV. A digitális képek felhasználásával kaptuk AMT Imaging System (Advanced Techniques Mikroszkóp Corp, Danvers, MA). Katalógusa Eredmények katalógusa génexpressziós aláírás gyomorrák katalógusa Az 1. táblázat jellemzőit a betegeket Yonsei adatok [17]. Gyomorrák többnyire található a gyomor distalis és színpadi III /IV. A génexpresszió microarray adatok azon betegek, azt találtuk, 3360 tumor-specifikus géneket, melynek átlagos expressziós intenzitásokat megváltoztatták legalább két-szerese jelent normális szövet génexpresszió ( p a potenciális célzó gyógyszerek gyomorrák, a gyomorrák adott aláírás, mint bemenő lekérdezést kapcsolati térkép leírt "módszer" részben. Mi kifejezetten nézett vegyületek, hogy volt egy aláírás fordítottan korrelált a gyomorrák-specifikus aláírás és az azonosított különböző drogok, amelyek a 2. táblázat összegzi A rangsor jelölt szerek alapozva hozták létre inverz korreláció értéke és a p-érték. 2. táblázat (oszlopok 2-4) mutatja a legmagasabb rangú vegyületek Yonsei adatokat. Kapcsolódás térkép elemzés kimutatta, hogy a hiszton deacetiláz (HDAC) gátló, köztük vorinostatot és trichostatin A jelentenek potenciális jelöltek célzó gyomorrák. A foszfatidil-inozitol-3-kináz inhibitor LY294002, fenotiazin trifluoperazin, és a hő-sokk fehérje inhibitor tanespimycin is azonosítottak, mint jelölt cél szerek gyomorrák. Következő, validált eredményeink felhasználásával egy független halmaza génexpressziós adatok Stanford microarray Database (2. táblázat; Stanford adatok; oszlopok 5-7). Kapcsolódás térkép elemzése ezt az adathalmazt megerősítette vorinostat mint a legmagasabb rangú jelöltet. Összefoglalva, a Kapcsolatok Map elemzés azonosított vorinostat potenciális terápiás szer gyomorrák. Katalógusa Vorinostat mutatnak terápiás hatékonyságot in vitro katalógusa gyomorrákban sejtvonalak Hogy értékelje a terápiás hatásosságát vorinostat, megvizsgáltuk a növekedés a létrehozott gyomor rákos sejtvonalak (AGS, KATO-III és NCI-N87), miután 72 óra Vorinostat kezelés segítségével MTT assay. Összehasonlítva a kezeletlen sejtek, Vorinostat szignifikánsan gátolta a sejtek életképességét dózis-függő módon az összes gyomor rákos sejtvonalak (1a ábra). Igazoltuk a csökkentés a sejtek életképességét sejtciklus analízise AGS és KATO-III rákos sejteket. Kimutattuk, hogy a kezelés Vorinostat (5 uM) 24 órán indukált jelentős növekedést a sub-G1 aránya AGS sejtek, mint a kontroll (2,3 ± 0,07% vs 39,2 ± 0,99% -kal; P Összefoglalva, ezek az adatok azt jelzik, hogy Vorinostat rendelkezik antiproliferatív hatással gyomorrák sejtvonalak által apoptózis indukálására AGS sejtek és a G2 /M sejtciklus KATO-III-sejtekben. a globális génexpressziós analízise gyomor rákos sejtvonalak AGS és KATO-III után Vorinostat kezelés Ahhoz, hogy elemezzük a hatás Vorinostat globális génexpressziós, AGS és KATO-III gyomorrák sejteket kezeltünk 5 jiM Vorinostat 48 órán át, és microarray analízist végeztünk. Felügyelet nélkül klaszteranalízise microarray adatok után Vorinostat kezelés azt mutatta, hogy az AGS és KATO-III sejtek csoportosulnak az azonos sejtvonalból tekintet Vorinostat kezelés (2A ábra). Mivel autofágiát leírták, hogy szerepe van Vorinostat-indukált hatások más rákos megbetegedések [20], [21], végeztünk felügyelt elemzését autofágia kapcsolatos gén halmaz (80 gének és 149 szondák; táblázat S2). Vorinostat kezelt gyomorrák sejtvonalat egy csoportban (2b ábra), jelezve, hogy az indukciós autofágia fontos tulajdonsága Vorinostat gyomorrákban vonalak. Tekintettel arra, hogy az autofágia szerepe lehet mind a rákos sejtek túlélését és a rák sejthalál hatóanyaggal végzett kezelés után [22], [23], azt tovább értékeltük a hozzájárulás ez a folyamat a hatását Vorinostat a gyomor rákos sejtvonalak. Mi funkcionálisan értékelte ezt a folyamatot immunfoltelemzéssel az autofágia marker mikrotubulus-asszociált protein 1 könnyű lánc 3 (LC3). Vorinostat kezelt KATO-III sejtek, és kisebb mértékben AGS sejtek mutatott egyértelmű felhalmozódása a gyorsabb-migráló lipidált formája LC3 (LC3-II) (3A ábra). Felhalmozódása LC3-II következménye lehet túlszabályzását autofagoszóma képződés vagy elzáródása autofág lebomlása LC3-II [24], [25]. Ezért elemeztük Vorinostat kezelt sejtek P62 degradáció, a marker autofág fluxus [24], és azt tapasztaltuk, csökken a P62 fehérje szint Vorinostat kezelt sejtekben, mint az idő kiegyenlített kezeletlen rákos sejtek (3A ábra). Ezen túlmenően, cotreatment az Vorinostat a bafilomycin A1 (BafA1), inhibitor autofagoszóma-lizoszóma fúzió, tovább növelte LC3-II felhalmozódás (3B), kompatibilis a Vorinostat indukáló autofág fluxus helyett blokkolja a lebontó képessége autophagolysomes. Elektronmikroszkópos (EM) egy érzékeny, mennyiségi és végleges módszert az autofágia [24]. Összhangban a Western blot adatok, EM elemzés azt mutatta, hogy Vorinostat jelentősen nőtt autofagoszóma képződés AGS sejtekben (4b ábra és B '), mint a kontroll sejtek (4A, ábra, és A'). Annak vizsgálatára, hogy a felhalmozódása autophagosomes védi a sejteket a celluláris stressz által kiváltott vorinostat, mi gátolta a autofágia folyamatot a farmakológiai inhibitor klorokin, kis interferáló RNS (siRNS) ellen Beclin-1. Hozzáadása klorokvin, hogy Vorinostat kezelt AGS és KATO-III sejtek eredményezett dózisfüggő csökkenése életképesség (5A ábra). A csökkenés életképesség megerősítette KATO-III sejtek kezelt Beclin-1 siRNS (5B). Együtt, az adatok arra utalnak, hogy a gátlása Vorinostat-indukált autofágia javíthatja a hatékonyságát Vorinostat kezelésére gyomor rákos sejteket. Ahhoz, hogy megértsük a hatásait Vorinostat a génexpresszió gyomor rákos sejtvonalak, végeztünk microarray analízise Vorinostat kezelt AGS és KATO-III gyomorrák sejtvonalakon (ábra. 2). Az elemzés szignifikáns genetikai különbségek kezeletlen és kezelt vorinostat gyomorrák sejtek (AGS és KATO-III). Miután vorinostat kezelés, a kifejezés a 1014 gének nőtt, és a kifejezés a 760 gén csökkent az AGS sejtvonalban. Kato-III sejtvonal 164 gének voltak, és 191 géneket szabályozva (kétszeres különbség; P katalógusa < 0,001). Vorinostat jelentősen megváltozott a kifejezés a 140 gének egyaránt AGS és KATO-III sejtvonalak (vorinostat specifikus gén aláírás). A gének, amelyek a leggyakrabban után megváltoztatták vorinostat kezelés azonosítottak SPANXA1, SPANXA2, VGF, DHRS2, ENTPD8, PNPLA7, STX1A, ARRDC4, KRT13, PRPH, NEU1, TXNIP, CCK katalógusa (> 4-felhajtható -regulation) és a MUC1, IFITM1, ANKRD37 katalógusa (> 4-szeres down-reguláció). katalógusa Ezután azt hivatott azonosítani biomarker jelöltek előre jelezni képes vorinostat érzékenysége az emberi gyomorrákos betegek . Ezért kombinált halmazát megváltozott gének Vorinostat kezelt gyomor rákos sejtvonalat (Vorinostat specifikus gén aláírás), és a két emberi gyomorrák aláírást generálunk a Yosei és a Stanford adatokat Venn-diagram elemzés. Azt találtuk, hogy a relatív expressziós szint 12-géneket megfordítható Vorinostat kezelés (6. ábra). Ezek közül 12 gén, 7 gének erősen kifejezett gyomorrák szöveteket lecsökkent ( ITGB5 katalógusa, TYMS katalógusa, MYB katalógusa, APOC1 katalógusa, CBX5 katalógusa, PLA2G2A katalógusa, KIF20A katalógusa), és 5 alacsony expresszált gének voltak akár szabályozott után vorinostat kezelés ( SCGB2A1 katalógusa, TCN1 katalógusa, CFD katalógusa, APLP1 katalógusa, NQO1 katalógusa (3. táblázat). katalógusa Vita katalógusa Eredményeink azt mutatják, hogy a globális humán gyomorrák gén aláírás hasznos lehet találni terápiás szerek, amelyek inkább a cél a genomiális aláírását gyomorrák megcélzása helyett egy vagy két specifikus gének. a kapcsolati térkép, azt találtuk, hogy a HDAC-inhibitorok, mint például a Vorinostat és Trichostatin a, kellett egy fordított korrelációban gén aláírás összehasonlítva a gyomorrák specifikus gén aláírás, és ezért is lehet ólom terápiás jelöltek gyomorrák. in vitro Eredményeink azt mutatták, hogy a HDAC inhibitorok, Vorinostat és trichostatin a, volt az első terápiás jelöltek gyomorrák, amely egyetért a koncepció, hogy a HDAC overexpresszálódik gyomorrákban szövetekben [26]. HDAC inhibitorokról kimutatták, hogy növelje acetiláció hisztonok, ezért érintő génexpressziót. Ezek az inhibitorok nyilvánvaló rákellenes hatása által indukálva az intrinsic és extrinsic apoptózis útvonal [27], [28], blokkoló tumor angiogenezis [29], és gátolja az intracelluláris stresszválasz utakat [30]. Mivel a HDAC inhibitorok átfogó hatása a génexpresszió, ezek befolyásolhatják a még feltáratlan sejtes folyamatokat [31], [32]. Klinikai beállítások HDAC inhibitorokat többnyire alkalmazott malignus hematológiai betegségben, de a klinikai vizsgálatok során a szilárd tumorok folyamatban vannak. Egy nemrégiben készült tanulmány, amely támogatja a in vitro katalógusa eredmények azt mutatták, terápiás hatékonyságát HDAC inhibitorok humán gyomorrák minták felhasználásával histoculture terápiás válasz vizsgálat [33]. Ugyanakkor azt is meg kell-e feltáratlan specifikus molekuláris meghatározott alcsoportok megjósolni választ, vagy ellenállás HDAC inhibitorok. Ezen kívül az alkalmazás potenciálisan hasznos biomarkerek még számos korlátja van [34]. Katalógusa A Connectivity térkép továbbra is nehéz felmérni, amíg nem tudjuk felmérni, hogy milyen mértékben genomi aláírásokat nyert in vitro katalógusa kísérletek összefoglalja a bonyolult emberi betegség. Bár a kapcsolati térkép több mint 7000 expressziós profilok képviselő 1309-vegyületek, a megközelítése is foglalkozik bizonyos korlátozások, mint például a korlátozott mennyiségű sejtvonal adatok (MCF7, PC3, stb), és a tudatlanság mikrokörnyezet befolyása az emberi test [16] . Ezen túlmenően, génexpressziós profilok származó kezelésére tenyésztett humán sejtek esetleg nem korrelál a in vivo In vitro katalógusa értékelése terápiás hatékonyságát vorinostat a gyomor rákos sejtvonalak kiderült, hogy ez a terápiás hatóanyag, kimutatták, antiproliferatív hatás fiziológiás releváns dózisok (5 uM) kompatibilis más publikációk: IC 50 AGS és KATO-III kimutatták, hogy rendre 2,9 uM és 5,9 jiM [ ,,,0],35]. Elemzés a sejtciklus (1B ábra), a sejtek életképességét és (1C) apoptotikus válaszok (1D ábra) azt mutatja, hogy a hatás a Vorinostat eltérést mutatnak a különböző gyomor rákos sejtvonalak, apoptózis indukálására AGS sejtek és a G2 /M letartóztatás Kato -III sejtek, ill. A különbség a genetikai /mutációs háttérben a sejtvonalak lehet felelős az eltérést. Microarray génexpressziós analízis összehasonlításával Vorinostat kezelt és a kezeletlen gyomor rákos sejtvonalakban jelezte, hogy ez a gyógyszer által indukált autofágia, amely összhangban van azzal a korábbi jelentések más rákos sejtvonalakban [20], [21]. Mivel mind indukció és a gátlás autofágia lehet terápiás előnyöket [22], [23], mi is értékeltük a szerepét autofágia gyomorrák sejtek után vorinostat kezelést. Gátlása autofágiát segítségével egy jól ismert maláriaellenes szer, a klorokvin [21] és használata elleni siRNS Beclin-1 csökkenését eredményezte az életképességét gyomor rákos sejtvonalak jelenlétében vorinostat, ami arra utal autofágiának aktiválódik, mint egy védő túlélési válasz után Vorinostat kezelés (5. ábra). Ezáltal kombinálva rákellenes szerek, mint vorinostat gátló és autofágia nyújthat terápiás előny elleni küzdelemben gyomorrák. Katalógusa azonosítására vorinostat indukált gén aláírás értékeltük a génexpressziós profilokat AGS és KATO-III sejt sorok után vorinostat kezelést. A kifejezés a 1774 és 355 gén fordított volt (> 2-szeres, p < 0,001) AGS és KATO-III-sejtvonalak, ill. Ez arra utal, hogy az éves növekedési sejtvonal sérülékenyebb, mint a KATO-III vorinostat kezelés tekintetében génexpresszió. Ez a jelenség magyarázhatja az érzékenységet a AGS sejtek vorinostat. Felfedeztük, hogy Vorinostat jelentősen megváltoztatta a kifejezés egy sor 140 gének egyaránt AGS és KATO-III-sejtvonalakban.
és a KIF20A
) amelynek expressziója emelkedett volt gyomor tumorszövet és downregulált több, mint 2-szeresére Vorinostat kezelés gyomor rákos sejtvonalak. Ezzel szemben, a SCGB2A1, TCN1, CFD, APLP1, katalógusa, és NQO1 katalógusa nyilvánul fordított mintát. Katalógusa
alkalmazásával kapcsolati térkép elemzés azonosítani terápiás szerek, amelyek potenciálisan ellen hatásos ez a fajta rák. Mi is alátámasztotta a felső célzó gyógyszer hatásosságát a gyomor rákos sejtvonalak. Azt találtuk, hogy vorinostat, mint potenciális új gyógyszer, indukált apoptózist és autofágia gyomor rákos sejtekben. Együtt, ez a vizsgálat azt mutatja, hogy a kapcsolati térkép analízis lehet azonosítására használt terápiás szerek, amelyek lehetnek sikeres a kezelés egy részhalmaza gyomor rák.
Módszerek
A sejt növekedés, az életképesség és a sejtciklus esszék
Western-blot-analízis
siRNS transzfekció
transzmissziós elektronmikroszkópos
< 0,001, ábra S1 ). Ez az 3360-gének (azaz a gyomorrák-specifikus aláírás) használtuk a további in silico
szűrés lehetséges terápiás hatóanyagok gyomorrák.
kapcsolati térkép analízis megállapítja potenciális célzó gyógyszerek gyomorrák
<0,01), jelezve az indukciós a sejthalál (1B ábra). Ezzel szemben a sub-G1 aránya Vorinostat kezelt KATO-III sejtek nem befolyásolta (1B ábra), míg az arány a G2 /M sejtek jelentősen nőtt (21,4 ± 1,91% vs. 29,3 ± 0,35%; P
= 0,044), utalhat sejtciklusmegállás. Mi tovább teszteljük a sejtek életképességét a PI-kizárás. Kezeltük AGS és KATO-III sejtek 5 nM vorinostat 72 óra és értékelni őket áramlási citometriával (1C). Az összeg az elhalt sejtek, amelyek alacsony forward scatter és magas oldali szórás, szignifikánsan emelkedett AGS sejtekben (12,4 ± 4,3% vs. 79,4 ± 5,7%), és szintén a KATO-III sejtek, kontroll sejtekhez viszonyítva (8,7 ± 0,6% vs. 46,8 ± 2,1%). Mi végül elemeztük az apoptózis indukcióját követően Vorinostat kezelés AGS és KATO-III gyomorrák sejt vonalak segítségével immunoblot. Vorinostat fokozott apoptózist, értékelése szerint kaszpáz-3-hasítás, a AGS sejtek és kisebb mértékben Kato III sejtek (1D ábra).
gátlása autofágia fokozza Vorinostat hatásosság gyomorrák sejtek katalógusa
Vorinostat változik gén aláírás humán gyomorrák sejtek
értékelése a terápiás hatékonyságát Vorinostat kiderült, hogy ez a terápiás gyógyszer elfojtott növekedése a különböző gyomorrák sejtvonalakban. Továbbá annak antiproliferatív hatást, vorinostatot is túlszabályozott autofágia-specifikus géneket. Gátlása vorinostat indukált autofágia eredményezett további csökkenését életképességét. Ezen kívül, kombinált elemzése gyomor rákos sejtvonalak kezelt Vorinostat és mintákat a gyomor rákos betegek azt mutatta, hogy Vorinostat megváltoztatta az expressziós szintek egy sor tizenkét gyomorrák specifikus géneket.
rákellenes hatás, mivel a komplex természete a rák. Ezen kívül, a bemeneti csak korlátozott mennyiségű gének megengedett, amely torzítás a kapott eredményeket. További vizsgálatok foglalkozni kell, hogy az általunk kapott eredmények révén CMAP elemzés megerősítette, egy in vivo katalógusa gyomorrák modell. Annak ellenére, hogy ezeket a korlátozásokat, kapcsolati térkép elemzése potenciálisan hasznos módszer az új funkciók gyógyszerek, beleértve vorinostat, már használatban lévő klinikán más célra. Katalógusa
 Metanogének:a puffadás oka, amiről még nem hallottál (és egy új megoldás a SIBO számára)
Metanogének:a puffadás oka, amiről még nem hallottál (és egy új megoldás a SIBO számára)
 Emberek versus vírusok - El tudjuk kerülni a kihalást a közeljövőben?
Emberek versus vírusok - El tudjuk kerülni a kihalást a közeljövőben?
 A vastagbélrák elkerülése
A vastagbélrák elkerülése
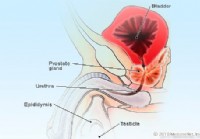 Prosztatagyulladás vs. BPH (megnagyobbodott prosztata):mi a különbség?
Prosztatagyulladás vs. BPH (megnagyobbodott prosztata):mi a különbség?
 Az immunterápiában részesülő betegeknek több rostot kell fogyasztaniuk,
Az immunterápiában részesülő betegeknek több rostot kell fogyasztaniuk,
 Az egyenes igazság az online egészségügyi üzlet felépítéséről
Az egyenes igazság az online egészségügyi üzlet felépítéséről
 Emésztési zavarok:vizuális útmutató a gyomorfekélyekhez
Mi az a fekély? Ha a gyomra egészséges, ragacsos nyálkaréteg borítja. Ez megvédi az ételt lebontó durva savtól. Ha valami felborítja ezt az egyensúlyt, akkor több gyomorsav és kevés nyálka keletkezi
Emésztési zavarok:vizuális útmutató a gyomorfekélyekhez
Mi az a fekély? Ha a gyomra egészséges, ragacsos nyálkaréteg borítja. Ez megvédi az ételt lebontó durva savtól. Ha valami felborítja ezt az egyensúlyt, akkor több gyomorsav és kevés nyálka keletkezi
 Tökéletes kakik (a rádióban)
izé! Nemrég értünk haza az Ancestral Health Symposiumról Atlantában, Georgia államban. Az egészségügyi konferenciák sokat kivehetnek belőled (jó értelemben). Rengeteg prezentációt is meg kell hallga
Tökéletes kakik (a rádióban)
izé! Nemrég értünk haza az Ancestral Health Symposiumról Atlantában, Georgia államban. Az egészségügyi konferenciák sokat kivehetnek belőled (jó értelemben). Rengeteg prezentációt is meg kell hallga
 Hogyan találjuk meg emésztési problémái kiváltó okát?
Ez a kép az éves újratesztelésemről. Tartalmazza a legjobb minőségű vérvizsgálatok, vizeletvizsgálatok, nyálvizsgálatok és székletvizsgálatok széles skáláját… Ó, óóó! Idén még 2057 dollárt hozott vis
Hogyan találjuk meg emésztési problémái kiváltó okát?
Ez a kép az éves újratesztelésemről. Tartalmazza a legjobb minőségű vérvizsgálatok, vizeletvizsgálatok, nyálvizsgálatok és székletvizsgálatok széles skáláját… Ó, óóó! Idén még 2057 dollárt hozott vis