abstraktné
Nesfatin-1 sa vylučuje, jedlo reagujúci anorexigenic peptid kódovaný v prekurzoru nucleobindin-2 [NUCB2]. Cirkulujúci nesfatin-1 zvyšuje post-prandially, ale diétne zložky, ktoré modulujú NUCB2 /nesfatin-1 zostávajú neznáme. Predpokladali sme, že sacharidov, tukov a bielkovín rozlične regulujú tkanivovo špecifické expresie nesfatin-1. NUCB2, prohormón konvertázy a nesfatin-1 boli zistené v myšiam žalúdočnej ghrelinoma [MGN3-1] bunkách. NUCB2 mRNA a proteín sa zistili aj v pečeni myší, a tenkého a hrubého čreva. MGN3-1 bunky boli ošetrené s glukózou, mastné kyseliny alebo aminokyseliny. Samce myší C57BL /6 boli dlhodobo kŕmené vysokým obsahom tuku, vysoký sacharidov a vysokým obsahom bielkovín stravy po dobu 17 týždňov. Kvantitatívne PCR a nesfatin-1 testy boli použité pre určenie nesfatin-1 na mRNA a proteínovej úrovni. Glukóza stimulovaná expresie mRNA v NUCB2 MGN3-1 bunkách. L-tryptofán tiež zvýšenie expresie NUCB2 mRNA a expresie mRNA ghrelinu a nesfatin-1 sekrécie. kyselina olejová inhibuje expresiu NUCB2 mRNA, zatiaľ čo expresia ghrelinu mRNA a sekréciu bola posilnená. NUCB2 mRNA expresie bol v pečeni myší významne nižšie kŕmené s vysokým obsahom bielkovín stravy v porovnaní s myšou kŕmených iné diéty. Chronického príjmu stravy s vysokým obsahom tuku spôsobil významnú redukciu NUCB2 mRNA v žalúdku, zatiaľ čo s vysokým obsahom bielkovín a vysokým obsahom tuku strava spôsobila podobné potlačenie NUCB2 mRNA v hrubom čreve. Žiadne rozdiely v sére nesfatin-1 hladiny boli nájdené u myší v 7 a.m, na začiatku fázy svetla. S vysokým obsahom sacharidov kŕmené myši vykazovali významne zvýšené nesfatin-1 hladiny v sére 1 hod nesfatin-1, bol významne nižší u myši kŕmené s vysokým obsahom tuku, proteín alebo sacharid porovnaní s kontrolami 7 p.m, tesne pred temné fáze. Myši, ktoré dostali bolus vysokým obsahom tuku sa významne zvýšené nesfatin-1 /NUCB2 u všetkých testovaných po žalúdočnej sonda časových bodoch, v porovnaní s kontrolnou myšou a myšou privádza ďalšie diéty. Naše výsledky prvýkrát ukazujú, že nesfatin-1 je modulovaný živinami
Citácia :. Mohan H, Ramesh N, Mortazavi S, Le A, Iwakura H, Unniappan S (2014) Živiny Diferenčný Regulovať Nucleobindin-2 /Nesfatin-1 in vitro Editor: Yvette Tache, University of California, Los Angeles, United States of America prijatá: 01.5.2014; Prijaté: 18.listopadu 2014; Uverejnené: 15 decembra 2014 Copyright: © 2014 Mohan et al. Toto je článok o otvorený prístup distribuovaný pod podmienkami Creative Commons Attribution licencie, ktorá umožňuje neobmedzené použitie, distribúciu a reprodukciu v nejakom médiu, za predpokladu, že pôvodný autor a zdroj sú pripísané Dostupné dát: Text. autori potvrdzujú, že všetky údaje súvisiace závery sú plne k dispozícii bez obmedzenia. Všetky relevantné údaje sú v papiera a jeho podporné informácie súbory Financovanie: Tento výskum je financovaný z otvoreného prevádzkovej dotáciu z kanadských Insitutes zdravotníckeho výskumu (CIHR http :. //www.cihr-irsc. gc.ca/e/193.html) a prevádzkarne Grant od Saskatchewan Health Research Foundation (http://shrf.ca/). SU je CIHR New vyšetrovateľ. Platcovia mal žiadnu úlohu v dizajne štúdie, zber a analýzu dát, rozhodnutie publikovať, alebo príprave rukopisu. Investori nemajú žiadnu úlohu vo výskume alebo rukopis Konflikt záujmov: .. Autori vyhlásili, že žiadne konkurenčné záujmy neexistujú Úvod Nesfatin-1 [NEFA /NUCB2 -encoded sýtosti a tuku ovplyvňuje proteín-1] je účinný anorexigenic peptid podieľa na regulácii energetickej bilancie a homeostázy glukózy [1], [30]. To je kyselina 82 amino peptid odvodený z prekurzorového proteínu, nucleobindin-2 (NUCB2) [1]. NUCB2 sa skladá z 396 aminokyselín, ktorý sa skladá z dvoch ručných motívov EF a väzbové doména DNA [1], [3]. Post-translačný spracovanie podľa prohormón konvertázy (PC 1/3 a PC 2) spôsobí, že NUCB2 sa štiepi na tri peptidy, nesfatin-1 (1-82 aminokyselín), nesfatin-2 (85-163 aminokyselín), a nesfatin- 3 (166-396 aminokyselín). NUCB2 /nesfatin-1 sekvencia aminokyselín je vysoko konzervatívny naprieč stavovcov [6], [7], [31]. NUCB2 /nesfatin-1 sa vyskytuje v rôznych hypotalamu jadier, ktoré sú zapojené do energetického metabolizmu, ako nucleus arcuatus, paraventrikulárním jadru, jadru supraoptic, bočné hypotalame oblasti a zona increta [8], [9]. produkujúce inzulín beta bunky ko-expresný nesfatin-1 v ostrovčekoch pankreasu krýs a myší [2], [4], [11], čo naznačuje, že nesfatin-1 by mohol zohrávať dôležitú úlohu v sekréciu inzulínu a homeostázy glukózy [4], [29]. Ghrelinu a NUCB /nesfatin-1 sú colocalized v žalúdočných oxyntických slizničných žliaz u hlodavcov [13], a ľudia [16]. Bolo zistené, NUCB2 expresie mRNA v purifikovaných žalúdočnej sliznice endokrinných buniek je vyšší ako v mozgu krýs [13]. Po celej dĺžke NUCB2 proteín bol pozorovaný v tenkom a hrubom čreve a pečeni samčích krýs a myší ICR [5]. Široká distribúcia NUCB2 /nesfatin-1 v centrálnych a periférnych tkanivách ukazuje na úlohu nesfatin-1 pri regulácii metabolizmu. Denné podávanie nesfatin-1 spôsobilo rozšírené zníženie príjmu potravy a telesnej hmotnosti [1] , Intracerebroventrikulární podávanie NUCB2 potláča príjem potravy, telesnej hmotnosti a podkožné, mezenterické a hmotnosť tvorenie tuku u dospelých zvierat v závislosti na dávke. Okrem toho, NUCB2 knockdown v krýs infúzii antisencie oligonukleotid (ako morfolínovom-MON) spôsobila zvýšenie chuti do jedla a telesnej hmotnosti [1]. Intra-Paraventrikulární jadro injekcie nesfatin-1 znižuje kumulatívne príjem potravy na 1 až 3 hodiny [9]. Intraperitoneálnej injekcie nesfatin-1 malo za následok zníženie príjmu potravy u odolné proti leptínu db /db myší a Nesfatin-1 je jedlo citlivý glucoregulatory hormón [12], [13], a pankreatických ostrovčekov potkanov uvoľnenie NUCB2 v odpovedi na glukózu [14]. V klinických štúdiách, glukózy ošetrené predmety mali vyššie bazálnej nesfatin-1 úrovne v porovnaní s kontrolnými orgánmi [10]. V min6 bunkách, bolo pozorované zvýšenie 4-krát v nesfatin-1 hladiny, keď boli bunky inkubované v vysokým obsahom glukózy (16,7 mM) v porovnaní s nízkou glukózy (2,0 mM) [4]. Nesfatin-1 zvýšená glukóza stimulovanú sekréciu inzulínu z kultivovaných min6 buniek, ktoré boli inkubované v vysokým obsahom glukózy než pri nízkej glukózy v závislosti na dávke [4]. V slinivke streptozotocínom (STZ) -injected myší s diabetom typu 1, bolo zistené, že ako NUCB2 a preproinzulin Expresia mRNA boli významne nižšie [4]. Naproti tomu bolo zistené, že zvýšená nesfatin-1 kolokalizace s inzulínom v ostrovček beta buniek s vysokým obsahom tuku vyvolané obéznych myší s diabetu 2. typu. Nesfatin-1 má tkanivovo špecifické účinky na vychytávanie glukózy v krysích adipocytoch a svalu [30]. Celkovo možno povedať, nesfatin-1 vykazuje významnú úlohu v regulácii celé telo glukózu a energetickej homeostázy. Kým nesfatin-1 sa objavuje ako dôležitý jedlo reagujúci peptidu [1], [12], [13], [30] , čo spúšťa jeho sekrécia zostáva nejasný. Aké súčasti stravy spustiť po jedle sekréciu nesfatin-1? Táto otázka zostáva nevyriešených. Hlavným cieľom tejto štúdie je zistiť, ako rôzne živiny môžu modulovať bunky NUCB2 /nesfatin-1 in vitro Ethics Prehlásenie Všetko štúdie využívajúce zvieratá splnené kanadskej Rady pokynov starostlivosti o zvieratá, a boli schválené Animal Research Ethics rady University of Saskatchewan (Protocol Number 2012 - 0033). in vitro štúdií myši žalúdka ghrelinoma (MGN3-1) bunky [17] boli kultivované v DMEM (Invitrogen, Ontario, Kanada, katalógw95-040), ktoré bolo doplnené 10% fetálnym hovädzím sérom (Invitrogen, katalóg|84 ) a 1% penicilínom (100 u /ml) a streptomycínom (100 ug /ml) (Invitrogen, katalóg—40-122) pri 37 ° C v 10% CO 2. Pri 80% zhlukovania, MGN3-1 bunky boli schovať na 6 x 10 6 buniek /jamku v 12-jamkové doštičky a boli vykonané štúdie, kedy boli bunky 80 až 90% zhlukovania. Každá štúdia bol opakovaný trikrát a dáta z troch štúdií boli spojené za získania n = 9-12 jamkách /ošetrenie. Na určenie, či glukózy mal účinok v dávke a na čase závislým spôsobom, bunky boli inkubované po dobu 1 hodiny a 2 hodiny s 5,6, 25, 50, a 100 mM glukózy média DMEM. Úplné rastové médium z MGN3-1 buniek vyžaduje, aby boli rastúce na vysokej hladiny glukózy, čo je 25 mM. Vo vzťahu k štúdiách vykonaných s mastnými kyselinami a aminokyselín, sme vykonali tieto štúdie s použitím DMEM pri nízkych hladín glukózy (5,6 mM), pretože s použitím médium s vysokým obsahom glukózy (25 mM), by mohli maskovať účinok príslušných živín na NUCB2 /nesfatin -1 sekrécie a syntéza. Pokiaľ ide o mastné kyseliny s dlhým reťazcom, sme testovali vplyv troch rôznych mastných kyselín za použitia kyseliny linolenovej (Sigma-Aldrich, Ontario, Kanada; Product # L2376), kyselina oktánové (Sigma-Aldrich, Product # C2875), a kyselina olejová (Sigma -Aldrich; Product # O1383). Bunky boli inkubované po dobu 4 hodín za každý mastné kyseliny sa pri teplote 0, 1, 10, 100 uM. Použili sme L-tryptofán (Sigma-Aldrich, product # T8941) na testovanie účinku aminokyseliny na NUCB2 sekréciu a syntézu. Bunky boli inkubované počas 4 hodín s L-tryptofán na 0,7, 1, 10 mM. L-tryptofán je prítomný v kontrolnom médiu (5,6 mM glukózy v DMEM) pri minimálnej dávke 0,7 mm, ktorá je nevyhnutná pre ich stavu rastu. pre chronické podávania stravy, ktoré obsahujú rôzne množstvá konkrétneho živín, vek a hmotnosť-uzavreté (5 týždňov, priemerná telesná hmotnosť 20 gramov) mužské myší C57BL /6 (Charles River Laboratories, Quebec, Kanada) boli umiestnení jednotlivo pre 17 týždňov v 12 hodín svetla: 12 hodín tmy (vypínanie svetiel v 7 hodín a v v 7 hodín ráno), regulovanou teplotou a vlhkosťou terárium. Myši boli rozdelené do štyroch skupín kŕmených kontrolné (n = 6), vysokým obsahom sacharidov (n = 7), s vysokým obsahom bielkovín (n = 7), a s vysokým obsahom tuku (n = 7), diéta s ad libitum V akútnom podaní živín, veku a hmotnosti zhodou (5 týždňov staré, priemerné telesnej hmotnosti .: 20 gramov) mužské myší C57BL /6 (Charles River Laboratories, St Constant, QU, Canada) boli umiestnené jednotlivo na 1 týždeň a 2 dni v 12 h svetlo: 12 h tmy (svetla vypnúť v 7 hodín a v 7 hodín ráno ), riadenou teplotou a vlhkosťou terárium. Myši boli aklimatizovať počas 1 týždňa pri príchode a mal ad libitum prístup k vode a krmivu pre myši pravidelnej po dobu 11 dní. Pretože sme sa vykonáva akútne diétne štúdii sme potrebovali zvieratá aklimatizovať na orálny sondou konania. aklimatizáciu sme myší k tomuto postupu je gavaging vodovodné vodou po dobu 2 dní pred experimentálne deň. O 12 th deň boli myši na lačno po dobu 4 hodín a bola gavaged s určitou tekutú diétu. Myši boli rozdelené do 4 skupín: s vysokým obsahom bielkovín (Isopure proteínový nápoj, Zero Carb - Mango broskyňa príchuť; Nature je najlepšie, Clifton Park, New York; n = 7) s vysokým obsahom tukov (Splendido, za studena lisovaný extra panenského olivového oleja alebo prezidenta Choice , Canada; n = 7), s vysokým obsahom sacharidov (D-glukóza, BioShop; Catlogue # GLU501.500; n = 7), a vody (voda z vodovodu, n = 7). V deň štúdie, 200 mikrolitrov nad živín /voda bol podávaný myší orálny sondou. Hodnoty glukózy v krvi odfotil na 0, 5, 10, 15, 20, 30, 60, 90 a 120 minút, a bola odobratá krv na 15, 30, 60 a 120 minút pre analýzu ELISA pre stanovenie cirkulujúcich hladín NUCB2 /nesfatin -1. Tkanivá (žalúdok, tenké črevo [dvanástnika], hrubého čreva a pečene) boli odobraté z každej myši po ukončení štúdie (hlboko izoflurán eutanáziu a následne cervikálny dislokáciou). Aby bola zachovaná konzistencia, časovanie a trvanie každého pokusu, operácie a odber vzoriek boli udržiavané konštantné pre všetky štúdie. Bunky alebo tkanivá boli odobraté z každej štúdie porovnať expresie NUCB2 mRNA. Z myší, ktoré podstúpil testovanie v rámci chronickej diéta štúdiu, tkaniva (žalúdka, tenkého čreva, hrubého čreva a pečeň) boli zozbierané ihneď po eutanáziu. Celková RNA bola extrahovaná z buniek a tkanív MGN3-1, pomocou izolácie činidla TRIzolu RNA (Invitrogen). RNA čistota bola overená pomocou optickej hustoty (OD) absorpčná pomer (OD 260 nm /OD 280 nm) za použitia Nanodrop 2000C (Thermo, Vantaa, Fínsko). Iba vzorky s absorpčným pomerom väčším ako 1,8 bola použitá pre syntézu cDNA. Syntéza cDNA bola vykonávaná pomocou kitu pre syntézu cDNA iScript podľa pokynov výrobcu (BIORADE, Kanada). RT-PCR a QRT-PCR NUCB2, ghrelin, a RT-PCR pre PC 1/3 a PC 2 boli vykonané podľa podmienok uvedených v tabuľke 1, pomocou real-time PCR Detection System CFX Connect (Bio-Rad). Pre analýzu QRT-PCR mRNA expresie NUCB2 bola normalizovaná pomocou beta-aktínu ako gén housekeeping. PCR produkty pre NUCB2 v žalúdku, pečene a hrubého čreva a týchto génov (NUCB2, ghrelin, PC a PC 1/3 2) v bunkách MGN3-1 boli podrobené elektroforéze v 1% agarózovom géle za účelom overenia prepisy zosilnený. na predchádzajúce štúdie [4] na báze, sme použili beta-aktínu ako vnútorná kontrola pre normalizáciu signálu NUCB2 mRNA. Pri použití celkovej RNA, kde RNA kvantifikácie boli veľmi presné, kritické prahy pre beta-aktínu nevykazovali žiadnu variabilitu. Relatívna expresia mRNA NUCB2 je normalizované s beta-aktínu z tej istej vzorky v súlade s metódou Livak [31]. MGN3-1 bunky boli kultivované v prezentácii systéme Labtek komory (Nalge Nunc International, Rochester, NY) a boli ponechané rásť do blízkej zhlukovania. Bunky boli premyté 1X roztok fosfátového pufra (PBS, 2 x 5 minút, 25 ° C) a pevná v roztoku (PFA) v 1X PBS po dobu 10 minút 4% paraformaldehydom v pri teplote 25 ° C, nasleduje ďalšie premytie 1X PBS ( 3 x 5 minút, 25 ° C). Fixované bunky boli permeabilizovány v roztoku 0,3% Triton-X (Bioshop, Burlington, Ontario, Kanada) v 1X PBS po dobu 5 minút pri teplote miestnosti. Sklíčka boli inkubované v blokovacím pufri, obsahujúcom 10% kozie sérum v 1X PBS po dobu 1 hodiny pri teplote miestnosti. Bunky potom boli inkubované v primárnej protilátke (tabuľka 2) pri teplote 4 ° C cez noc. Sklíčka sa premyjú PBS (3 x 5 minút, 25 ° C) a inkubuje so sekundárnou protilátkou (tabuľka 2; protilátka PC1 /3 bol veľkorysý dar od Dr. Iris Lindberg, University of Maryland School of Medicine) po dobu 4 hodín pri teplote izbová teplota. Nakoniec boli rezy premyté PBS (3 x 5 minút, 25 ° C) a upevnené pomocou Vectashield montážnej médium obsahujúce 4 ', 6-diamidínov-2-fenylindolu (DAPI) (Vector Laboratories, Burlington, Ontario, Kanada). Bunky boli považované za použitia Nikon Eclipse Tí inverzného fluorescenčného mikroskopu (Nikon, Mississauga, Ontario, Kanada) a snímky boli nasnímané Nikon DS-Qi1 MC fotoaparát (Nikon). Snímky boli analyzované pomocou NIS-Elements základného výskumu softvér (Nikon) na Dell HP Workstation. Uvedené obrázky sú reprezentatívnymi bunky zafarbené ghrelinu, NUCB2, PC 1/3 a PC2. Pri vysokom rozlíšení zobrazenie, bunky boli zobrazené, analyzované a obrazy snímané Leica TCS SP5 konfokálního mikroskopu. pre potvrdenie prítomnosti nucleobindin-2 (NUCB2) v črevách a pečeni, tri, 3 mesiace boli použité staré C57BL /6 samce myší. Stručne povedané, pečeň a tenké a hrubé črevo sa získali a zvlášť pre Western analýzu alebo imunohistochémia. Tkanivá pre westernovým prenosom homogenizuje v T-per bielkovín tkanív extrakčný činidlom (Thermo Scientific,̑10), nasledovaný bielkovín stanovenie koncentrácie Bradford testu. Vzorky boli pripravené v 1X Laemmliho pufri s obsahom 0,2% 2-mercaptoetanolu (Bio-Rad,¡-0737 a -0710) a následne sa varí pri teplote 95 ° C po dobu 5 minút a následne na rozvíri. Celý objem vzorky (20 ul), z ktorých každá obsahuje 50 ug proteínu alebo syntetické potkan nesfatin-1 (ABGENT, 1 ug /ul, skôr používané v bode 4, 29) sa nanáša na gél, a spustiť v Mini-Protean TGX 8-16 % gradientom gél (Bio-Rad,Lj-1104). Po oddelení boli proteíny prenesené na 0,2 um BioTrace nitrocelulózové membránu (PALL Life Sciences,đ77-000), a potom bola membrána blokovaná v 1X RapidBlock roztoku (Amresco, # M325). Detekcia NUCB2 proteín bola vykonaná s použitím králičie anti-nesfatin-1 (Katalógové číslo H-003-22; 1: 500 riedenie, Phoenix Pharmaceuticals, Kalifornia) a GAPDH proteín bol detekovaný použitím králičím antisérom namierenej proti myšiemu GAPDH (AbDSerotec, # AHP1628 ) zriedený 1: 1000. Ako sekundárny protilátka, kozí anti-králičie IgG (H + L) HRP (Bio-Rad,ª-6515) zriedeného 1: 3000 bol použitý. Pre vizualizáciu proteínov bola membrána inkubovaná po dobu 5 minút v Clarity Western ECL substrát (Bio-Rad,ª-5061), a zobraziť pomocou ChemiDoc MP zobrazovacieho systému (Bio-Rad,ª- 8280) s detekciou chemiluminiscencie. Membrána odizolovanie medzi systémom detekcie proteínu bola vykonaná pomocou nástroja Obnovenie PLUS western blot stripping vyrovnávacej pamäte (Thermo Scientific,ǐ30). Presné a proteínové duálny XTRA štandardy (Bio-Rad,¡-0377) boli použité ako markery molekulovej hmotnosti. imunohistochemických štúdií sa tkanivá získané boli fixované v 4% formaldehydu po dobu 24 hodín pri teplote 4 ° C. Fixačné sa nahradí etanolom (tri 70% etanol), po ktorých nasleduje 10 minútach inkubácie pri teplote 4 ° C. Tkanivá potom boli uložené v 70% etanolu pri teplote 4 ° C a boli spracované a rozrezané na Prairie Diagnostic Services Inc. (PDS Inc., Western College of veterinárneho lekárstva, University of Saskatchewan). Parafínové rezy 4 um hrúbky boli pripravené pre imunobarvení. Tieto sekcie boli zbavené voskov s xylénom (inkubované dvakrát v 100% xylénu, 5 minút, 25 ° C) a rehydratované v odstupňované rade etanolu (dvakrát inkubujú v 100% etanolu a raz v každej 95% etanolu, 70% etanol, 50% etanol, 2 minúty každý, 25 ° C). Rezy boli potom inkubované s 3% peroxidu vodíka v destilovanej vode pre blokovanie endogénnej peroxidázy (30 minút pri izbovej teplote). Rezy potom boli blokované s proteín blok činidla bez séra (DAKO Corporation, Kalifornia, USA) po dobu 10 minút predtým, než je inkubované s primárnymi protilátkami. Tieto sekcie boli potom inkubované s králičie anti-nesfatin-1 (Katalógové číslo H-003-22; 1: 500 riedenie, Phoenix Pharmaceuticals, Kalifornia, USA) po dobu 24 hodín pri teplote miestnosti. Všetky snímky boli následne trikrát premyté 1 x PBS a inkubované s kozím anti-králičím IgG (Texas Red Red-Nesfatin-1; Katalógové číslo Ti-1000; 1: 100 zriedenie, Vector Laboratories, Kalifornia) sekundárnej protilátky po dobu 1 hodiny pri teplote miestnosti teplotu. Všetky primárne a sekundárne protilátky boli nariedia v riediacim činidlom protilátka (DakoCytomation, Mississauga, Ontario). Sklíčka sa trikrát premyjú 1 x PBS a sedemkrát sa destilovanou vodou. Nakoniec bola sklíčka osadené Vectashield médiom, ktoré obsahujú jadrový DAPI farbivá (modrá; Vector Laboratories, Burlingame, Kalifornia). Rezy boli videné v rámci Nikon Eclipse Ti-E obráteným fluorescenčným mikroskopom (Nikon Kanada, Mississauga, Kanada). Snímky boli nadobudnuté Nikon DS-Qi1 MC chladený monochromatickú kameru pripojenú k počítaču Dell HP Workstation a NIS prvkov základný výskum software pre spracovanie obrazu (Nikon Kanada, Mississauga, Kanada). Iba reprezentatívne obrazy tenkého a hrubého čreva farbenie pre NUCB2 /nesfatin-1 s DAPI sú uvedené v sekcii výsledky. Ak chcete skúmať závislé na živiny zmeny v NUCB2 /nesfatin-1 sekrécia z MGN3-1 buniek, médium bolo odňaté po konkrétnych inkubačnej doby. Aby sa zabránilo bunkové zvyšky boli vzorky centrifugovány (13000 otáčok za minútu po dobu 10 minút pri teplote 4 ° C) a horná 700 ul bolo skladované pri teplote -20 ° C až do nesfatin-1 meranie. Pre meranie v obehu NUCB2 /nesfatin-1, bola odobratá krv v 7 a.m (skoro po začiatku ľahká fáza), 1 hod (stred svetelné fáza) a v 7 p.m (pred začatím temné fázu). Vzorky krvi sa nechajú zraziť na ľade, a sérum sa oddelí odstredením (7000 otáčok za minútu po dobu 9 minút pri teplote 4 ° C) a skladované pri teplote -20 ° C, kým sa testy vykonané. NUCB2 úrovňou /Nesfatin-1 sekrécie v médiách boli merané pomocou Nesfatin-1 (1-82) (Rat) ELISA kit (Katalógové číslo EK-003-22, Phoenix Pharmaceuticals Inc., Kalifornia). Limit citlivosti testu bola 1,2 ng /ml pre nesfatin-1, s detegovateľným rozmedzí 0,1-1000 ng /ml. Množstvo imunoreaktívnych materiálu bola stanovená pomocou nelineárnej regresnej krivky-fit, ktorý bol použitý pre kvantifikáciu a porovnať koncentráciu NUCB2 /nesfatin-1 sekrécia vo vzorkách séra a médiá. Ak chcete skúmať živín závislé zmeny v NUCB2 /nesfatin-1 a celkové sekréciu ghrelinu z MGN3-1 buniek, médium bolo odňaté po konkrétnych inkubačnej doby. Aby sa zabránilo bunkové zvyšky boli vzorky centrifugovány (13000 otáčok za minútu po dobu 10 minút pri teplote 4 ° C) a horná 700 ul bol skladovaný pri teplote -20 ° C až do NUCB2 /Nesfatin-1 a celkové meranie ghrelinu. Vzorky krvi sa nechajú zraziť na ľade, a sérum sa oddelí odstredením (7000 otáčok za minútu po dobu 9 minút pri teplote 4 ° C) a skladované pri teplote -20 ° C, kým sa testy vykonané. NUCB2 /Nesfatin-1 úrovne sekrécie v sére a médiá boli merané pomocou Nesfatin-1 (1-82) (Rat) ELISA kit (Katalógové číslo EK-003-22, Phoenix Pharmaceuticals Inc., Kalifornia). Limit citlivosti testu bola 1,2 ng /ml pre nesfatin-1, s detegovateľným rozmedzí 0,1-1000 ng /ml. Rovnako tak celková úroveň sekrécie ghrelin v médiách bola meraná pomocou ghrelinu (potkan, myš) EIA kit (Katalógové číslo EK-031-31, Phoenix Pharmaceuticals Inc, Kalifornia). Limit citlivosti testu bola 1,16 ng /ml pre celkom ghrelinu, s detegovateľným rozmedzí 0-100 ng /ml. Množstvo imunoreaktívnych materiálu bola stanovená pomocou nelineárnej regresnej krivky-fit, ktorý bol použitý pre kvantifikáciu a porovnať koncentráciu NUCB2 /nesfatin-1 sekrécia vo vzorkách séra a médiá. Analýzy kvantifikovaných QRT-PCR a ELISA dát boli vykonané jednocestnú ANOVA nasledovanej Tukeyho viacnásobného porovnávacieho testu. GraphPad Prism verzia 5 (GraphPad Software Incorporated, San Diego, CA, USA) bol použitý pre štatistické analýzy a grafov. Význam bol pridelený, keď p menšie ako 0,05. Dáta sú vyjadrené ako priemer ± SEM. NUCB2, PC a PC 1/3 2 mRNA sú exprimované v bunkách a MGN3-1 NUCB2 mRNA je exprimovaný v žalúdku, pečene, malá črevo a hrubé črevo u myších samcov Zistili sme expresiu NUCB2 (202 bp), prohormón konvertázy 1/3 (400 bp), a prohormón konvertázy 2 (406 bp) mRNA v MGN3-1 buniek (obr. 1A). Expresia mRNA NUCB2 (202 bp) bol tiež detekovaný v žalúdku, pečene, tenkého a hrubého čreva samčích C57 /BL6 myší (Obr. 1B). Absolútna hladiny expresie mRNA NUCB2 v žalúdku boli vyššie než expresie NUCB2 mRNA v pečeni, tenkom čreve a hrubom čreve (viď obr. 1C). Fluorescenčné mikroskopia zobrazené MGN3-1 buniek zafarbených anti-nesfatin-1 protilátky (Texas-červená; obr. 2b) a anti-ghrelinu protilátky (FITC-zelená; obr. 2A) ukázala jasnú spoločnú lokalizáciu (žltá, obr. 2c) z nesfatin-1 a ghrelinu imunoreaktivity. Avšak, niektoré ghrelinu pozitívne bunky neboli imunoreaktívnych pre nesfatin-1 (obr. 2C). MGN3-1 bunky vykazovali PC 1/3 imunoreaktivitu (Texaská červeň, obr. 2D) a PC 2 imunoreaktivitu (Texaská červeň, obr 2E.). DAPI (modrá) zafarbené jadro všetkých buniek, vrátane tých, ktoré nie sú buniek pozitívnych na proteíny študovaných. Kontrolné sklíčka zafarbené samotným sekundárnou protilátkou (Obr. 2F) nemal imunoreaktivitu. Konfokálna zobrazovania ukázala MGN3-1 buniek zafarbených anti-nesfatin-1 protilátky (Texas-Red, obr. 3A) a anti-ghrelin protilátka (FITC-zelená; obr. 3B) ukázala jasnú spoločnú lokalizáciu (žltá, obr. 3C) z nesfatin-1 a ghrelinu imunoreaktivity. Negatívna kontrola je zafarbený sám iba sekundárne protilátky (obr. 3D). NUCB2 proteín je exprimovaný v hrubom čreve, tenké črevo a pečeň z myších samcov vykazujú zreteľný pás pre NUCB2 zodpovedajúce približne 50 kDa. Krysa nesfatin-1 peptid použitý ako pozitívna kontrola je zobrazený ako zreteľný pás, zodpovedajúci približne 10 kDa (obr 4A;. Ľavý obrázok). Avšak, žiadne pásy ukazujúce plne spracované nesfatin-1 boli viditeľné pri 10 kDa vo vzorkách tkanív (obr 4A;. Ľavý obrázok). Tiež sme zistili, pásma približne 47 kDa pod 50 kDa pásu v tenkom čreve a v pečeni, ale nie v hrubého čreva (obr 4A. Ľavý obrázok). Výrazným pásmo pre GAPDH použitá ako kontrolná domu k udržaniu génu je pozorovaná u 37 kDa je ukázané vo všetkých tkanivách (obr 4A;. Doprava obrazu). NUCB2 /nesfatin-1 imunoreaktivita je nájdený v slizničných buniek tenkého čreva (obr 4B. Ľavý obrázok) a hrubé črevo (obr 4B. Pravý obraz). bunky boli inkubované v 100 mM glukózy DMEM mal vyššiu expresiu mRNA NUCB2 než bunky boli inkubované pri 5,6, 25 a 50 mM koncentrácie glukózy v DMEM na 1 hodinu po inkubácii (obr. 5A). 2 hodiny po inkubácii sa bunky inkubujú v 100 mM DMEM boli významne vyššie u NUCB2 expresie mRNA než bunky boli inkubované pri 5,6 a 50 mM glukózy DMEM (obr. 5c). Po 1 hodine (Obr. 5B) a 2 hodiny (obr. 5D), neboli žiadne významné rozdiely v NUCB2 /nesfatin-1 sekrécie. NUCB2 mRNA expresie bol významne vyšší v bunkách inkubovaných pri 10 mM L-tryptofán (obr. 5E) v porovnaní s bunkami, inkubovaných pri 0,07 a 1,0 mM L-tryptofán. NUCB2 /nesfatin-1 sekrécia z bunky boli inkubované pri 1,0 a 10,0 mM L-tryptofán boli významne vyššie než bunky boli inkubované pri 0,7 mM L-tryptofán (obr. 5F). sme zistili, že expresia mRNA NUCB2 významne znížená u buniek ošetrených 1, 10 a 100 uM kyseliny olejovej (obr. 6e) v porovnaní s kontrolou. Žiadne zmeny v NUCB2 mRNA boli pozorované v bunkách ošetrených linolenová (obr. 6A) a kyseliny oktánové (Obr. 6C). Ďalej, NUCB2 /nesfatin-1 sekrécie bola nezmenená v bunkách liečených rôznymi dávkami linolénovej kyseliny (Obr. 6B), kyselina oktánové (obr. 6D), a kyseliny olejovej (obr. 6F). nemennosť ghrelinu mRNA (S1A obrázok) a celková sekréciou ghrelinu (obr S1B ) bolo pozorované, keď boli bunky ošetrené rôznymi dávkami glukózy po 1 hodine inkubácie. Expresia mRNA ghrelin bol významne vyšší v bunkách inkubovaných pri 1 mM L-tryptofán (obr. 7A), v porovnaní s bunkami, inkubovaných pri 0,07 a 10 mM L-tryptofán. Žiadny významný rozdiel v celkovom vylučovanie ghrelinu z buniek ošetrených rôznymi dávkami L-tryptofán (obr. 7B). Zistili sme žiadne zmeny v ghrelin expresie mRNA (Obr. 7C), ale celkové sekrécia ghrelin bol vysoký z buniek inkubovaných pri 100 uM kyseliny linolenovej (Obr. 7D) v porovnaní s kontrolou, 1 a 10 uM dávok. Žiadne zmeny v ghrelinu mRNA (obr. 7E) a celkového ghrelinu sekrécie (obr. 7F) bolo pozorované, keď boli bunky ošetrené rôznymi dávkami kyseliny oktánové. Medzitým, ghrelin mRNA (obr. 7H), a celkový ghrelin sekrécie (obr. 7I) boli významne vyššie v bunkách ošetrených s 100 uM kyselina olejová, v porovnaní s kontrolou, 1 a 10 uM kyseliny olejovej. telesnej hmotnosti, príjmu potravy a hladiny glukózy v krvi u myší kŕmených profil na rôznych diét sú znázornené na obrázku S2. Myši kŕmené stravou s vysokým obsahom tuku mali významne nízku expresiu NUCB2 mRNA v žalúdku v porovnaní s tými, privádza na ovládací prvok, vysokým obsahom bielkovín alebo vysokým sacharidov diéty (Obr. 8A). Nebol žiadny významný rozdiel v expresii mRNA NUCB2 v tenkom čreve medzi myšou kŕmených na ovládací prvok, s vysokým obsahom bielkovín, sacharidov s vysokou alebo vysokou diéty (obr. 8b). Myši kŕmené na vysokým obsahom bielkovín a vysoké diéty majú relatívne nízku expresiu NUCB2 mRNA v hrubom čreve, ako u myší kŕmených kontrolnej alebo s vysokým obsahom sacharidov diéty (obr. 8c). Myši kŕmené s vysokým obsahom bielkovín strava mala významný nízku expresiu NUCB2 mRNA v pečeni, než u myší kŕmených ostatné diéty (obr. 8D). 7 a.m, nedošlo v sére žiadne rozdiely nesfatin-1 /NUCB2 hladiny u myší kŕmených rôzne diéty (obr. 9a). 1 p.m, s vysokým obsahom sacharidov privádza myši mali významne vyššia v sére nesfatin-1 /NUCB2 v obehu (viď obr. 9B). Sérum nesfatin-1 /NUCB2 bola významne nižšia u myši kŕmené vysoké množstvo sacharidov, s vysokým obsahom bielkovín, alebo s vysokým obsahom tuku, v porovnaní s kontrolnými myšami napájané stravy v 7 hodín (obr. 9C). nedošlo k žiadnym významným zmenám v expresii mRNA NUCB2 v žalúdku (obr. 10A), tenké črevo (obr. 10B), hrubého čreva (obr . 10C) a pečene (viď obr. 10D) medzi myší kŕmených s vysokým obsahom bielkovín, vysokým obsahom sacharidov, s vysokým obsahom tuku alebo vody.
v kultivovaných žalúdku Ghrelinoma (MGN3-1) články a in Vivo
u myších samcov. PLoS ONE 9 (12): e115102. doi: 10,1371 /journal.pone.0115102
s vysokým obsahom tukov diétne privádza myšou [8]. Nesfatin-1 sa skladá z 3 štruktúrnych fragmentov a iba na polovici fragmentu (zvyšky 24-53; M30) z nesfatin-1 sa podieľa na tvorbe anorektické odpovedí [15], [28]. Spoločne tieto výsledky poskytujú jasný dôkaz, že podporujú sýtosti účinky nesfatin-1.
v kultivovaných žalúdku ghrelinoma (MGN3-1) z myší a in vivo
u mužov myši. Naše výsledky z našich in vitro
štúdií ukazujú, že MGN3-1 bunky reagujú odlišne na živiny v vylučovať NUCB2 /nesfatin-1 a ghrelinu. Rovnako tak akútny alebo chronický príjem živín robí vplyv NUCB2 expresiu mRNA a NUCB2 /nesfatin-1 uvoľňovanie určitým spôsobom diétu.
materiáloch a metódach
in vivo štúdií
prístup k vode a ich konkrétne diétu. Všetky diéty boli zakúpené od spoločnosti Research diets (New Brunswick, NJ). Obsah kalórií diét boli: kontrola (# výrobku D12451): 4,73 kcal /g s 20% energie získanej z bielkovín, 35% energie získanej zo sacharidov a 45% energie získanej z tukov; vysokým obsahom sacharidov (Product # D12450J) mal 3,8 kcal /g s 20% energie získanej z bielkovín, 70% energie získaná z sacharidov a 10% energie získanej z tukov; s vysokým obsahom bielkovín (Product # D08091802) mal 3,8 kcal /g s 60% energie získanej z bielkovín, 30% energie získaná z sacharidov a 10% energie získanej z tuku a vysokým obsahom tuku (Product # D12492) mal 5,2 kcal /g s 20% energia získaná z bielkovín, 20% energie získaná z sacharidov a 60% energie získanej z tukov. Všetky myši boli kŕmené s kontrolnou diétou počas jedného týždňa pred začiatkom ich špecifické stravy. príjmu potravy, telesná hmotnosť, hladina glukózy v krvi meraní pridanie 4 hodiny rýchlo sa merali raz týždenne po dobu 17 týždňov
Celková RNA extrakcia a syntéza cDNA
RT-PCR a kvantitatívnej real-time PCR
imunocytochémia a mikroskopia
Western blot analýza, imunohistochémia a fluorescenčné mikroskopia
Nesfatin-1 /NUCB2 v sére a médiá
NUCB2 /Nesfatin- 1 v sére a hladiny mediálnej a Total ghrelin v médiách
Štatistická analýza
Výsledky
MGN3-1 bunky sú immunopositive pre ghrelin, NUCB2 /Nesfatin-1 , PC 1/3 a PC 2
NUCB2 expresie proteínov v hrubého čreva, tenkého čreva a pečene od myších samcov
Vplyv glukózy a L-tryptofán na NUCB2 mRNA expresie v, a NUCB2 /nesfatin-1 sekrécie MGN3-1 buniek
Vplyv kyseliny linolenovej, kyseliny oktánové a kyseliny olejovej kyseliny na expresiu mRNA v NUCB2 a NUCB2 /nesfatin-1 sekrécie MGN3-1 buniek
Vplyv L-tryptofán, kyselina linolénová, kyselina oktánové a kyselina olejová nezávisle na expresiu mRNA ghrelin in, a celková sekrécia ghrelin z MGN3-1 buniek
Chronická účinky živín na NUCB2 expresie mRNA a sérum NUCB2 /nesfatin-1 u myší
Akútne účinky živín na expresiu mRNA NUCB2 , glukózy v krvi a sére NUCB2 /nesfatin-1 u myší
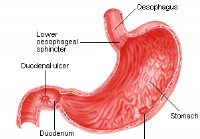 Peptické vredy
Čo je peptický vred? Peptický vred je lézia alebo vred na ochrannej výstelke žalúdka (kde sa nazýva žalúdočný vred) alebo dvanástnika. Lekári to často definujú ako prasknutie sliznice väčšie ako 3-5
Peptické vredy
Čo je peptický vred? Peptický vred je lézia alebo vred na ochrannej výstelke žalúdka (kde sa nazýva žalúdočný vred) alebo dvanástnika. Lekári to často definujú ako prasknutie sliznice väčšie ako 3-5
 Funkčné gastrointestinálne poruchy
Funkčné gastrointestinálne poruchy (FGD) sú poruchy tráviaceho systému, pri ktorých symptómy nemožno vysvetliť prítomnosťou štrukturálnych alebo tkanivových abnormalít. FGD nemajú identifikovateľné bi
Funkčné gastrointestinálne poruchy
Funkčné gastrointestinálne poruchy (FGD) sú poruchy tráviaceho systému, pri ktorých symptómy nemožno vysvetliť prítomnosťou štrukturálnych alebo tkanivových abnormalít. FGD nemajú identifikovateľné bi
 Čo spôsobuje vysoké hladiny kreatinínu?
Normálny rozsah hladín kreatinínu sa môže líšiť v závislosti od veku, pohlavia, veľkosti a svalovej hmoty osoby. Telo produkuje rôzne škodlivé chemikálie ako vedľajšie produkty metabolizmu. Kreatin
Čo spôsobuje vysoké hladiny kreatinínu?
Normálny rozsah hladín kreatinínu sa môže líšiť v závislosti od veku, pohlavia, veľkosti a svalovej hmoty osoby. Telo produkuje rôzne škodlivé chemikálie ako vedľajšie produkty metabolizmu. Kreatin