Background
kolinergisten tulehdusta reitti on sisäsyntyinen mekanismi, jonka autonomisen hermoston vaimentaa makrofagiaktivaatio kautta nikotiiniasetyylikoliinireseptorien (nAChR). Tämä käsite ei kuitenkaan ole osoitettu solutasolla koskemattomassa kudoksessa. Tätä varten olemme tutkineet vaikutusta nikotiinin aktivoinnista asuvan makrofagien hiiren vatsassa valmistelua avulla kalsiumin kuvantamisen.
Kalsium transientit ([Ca 2 +] i) kotimaisten makrofageissa kirjattiin hiiren vatsassa sisältävän valmisteen myenteeri- plexus ja lihasten kerrosta Fluo-4. Makrofagien aktivoituminen saavutettiin polttoväli pullistaa anto ATP. Vaikutukset nikotiinin makrofagien aktivoituminen arvioitiin ja nAChR mukana oli farmakologisesti ominaista. Läheisyys kolinergisten hermojen makrofagien kvantifioitiin konfokaalimikroskopialla. Expression of β2 ja α7 nAChR arvioitiin β2 immunohistokemiallisesti ja fluorofori-tagged α-Bungarotoksiini. 83% makrofagien kolinergisten suonikohjut hermosyiden havaittiin etäisyyksillä < 900nm. ATP aiheuttama [Ca 2 +] i lisätä estyi merkittävästi 65%: n tai 55% makrofaageihin 100gM tai 10gM nikotiinin, vastaavasti. Tämä estävä vaikutus on päinvastainen, jonka β2 nAChR mieluummin antagonisti dihydro-β-eryhtroidine mutta ei heksametoniumilla (ei-selektiivinen nAChR-antagonisti), mekamyyliamiinia (α3β4 nAChR-mieluummin antagonisti), α-Bungarotoksiini tai methyllycaconitine (sekä α7 nAChR-mieluummin antagonisti ). Makrofagit mahassa ilmaista β2 mutta ei α7 nAChR at proteiini tasolla, kun taas suolistossa ilmaista molemmat reseptorialayksiköitä. Tämä tutkimus on ensimmäinen Citation: Nemethova A, Michel K, Gomez-Pinilla PJ, Boeckxstaens GE, Schemann M (2013 ) Nikotiini Vaimentaa aktivointi Tissue Resident makrofagien hiiri Vatsa kautta β2 asetyylikoliinin nikotiinireseptorin. PLoS ONE 8 (11): e79264. doi: 10,1371 /journal.pone.0079264 Editor: Yvette Tache, University of California, Los Angeles, Yhdysvallat vastaanotettu: 10 kesäkuu 2013; Hyväksytty: 26 syyskuu 2013; Julkaistu: 01 marraskuu 2013 Copyright: © 2013 Nemethova et al. Tämä on avoin pääsy artikkeli jaettu ehdoilla Creative Commons Nimeä lisenssi, joka sallii rajoittamattoman käytön, jakelun ja lisääntymiselle millä tahansa välineellä edellyttäen, että alkuperäinen kirjoittaja ja lähde hyvitetään. Rahoitus: Tämä työ tukivat tukea Euroopan unionin seitsemäs puiteohjelma (ipodd), Deutsche Forschungsgemeinschaft Sche267 /9-1 MS; jonka myöntäminen Research Foundation Flanders (FWO, Odysseus ja Hercules-ohjelma) GEB, ja jonka FWO post doc Fellowship PJG. Rahoittajat ollut mitään roolia tutkimuksen suunnittelu, tiedonkeruu ja analyysi, päätös julkaista tai valmistamista käsikirjoituksen. Kilpailevat edut: Kirjoittajat ovat ilmoittaneet, etteivät ole kilpailevia intressejä ole. Vuonna 2000 Tracey ja työtoverit osoittivat, että sähköinen stimulaatio Kiertäjähermo tarjoaa voimakas tulehdusta tulo perna. Hiirimallissa sepsiksen, vagushermostimulaatiolle (VNS) alensivat pro-inflammatoristen sytokiinien tuotantoa, vaikutus riippuu α7 nikotiiniasetyylikoliinireseptorien (nAChr) [1,2]. Tämä niin kutsuttu "kolinergisen tulehdusta polku" (CAIP) toimii kautta adrenergisten pernan hermojen jolloin synaptisen kaltaisia yhteyksiä β2-adrenergisen reseptorin ilmentävien pernan T-soluissa. Myöhemmät asetyylikoliinin vapautumista (ACh) Näistä T-soluista vastaa tulehduksia oletettavasti vuorovaikutuksessa α7nAChR ilmentävien makrofagien [1,2]. Vuonna 2005 esittänyt todisteita siitä, että CAIP myös moduloi suoliston immuunijärjestelmään. Hiirimallissa postoperatiivisen suolentukkeuman, osoitimme, että VNS vähentää suolen manipulointi aiheuttama tulehdus ohutsuolen, suotuisa vaikutus riippuu α7 nAChR mutta riippumaton perna [3,4]. Nämä tiedot viittaavat siihen, että suolen immuunijärjestelmän melko suoraan moduloidaan vagaalisia hermopäätteitä ja /tai enteerisen neuronien. Koska asukas suoliston makrofagit ovat tärkeimmät toimijat käynnistävät tätä tulehdusreaktiota [5], nämä solut edustavat todennäköisimmin kohde CAIP. In vitro -tutkimukset eristetty peritoneaalimakrofageille, ääreisverenkierron mononukleaarisia soluperäisissä makrofagien tai makrofaagisolukantoja ovat todellakin täysin osoittaneet, että asetyylikoliinin ja nikotiini vähentää sytokiinien tuotantoa [1-3,6,7] ja lisätä fagosytoosin [6]. Kuitenkin on kyseenalaista, missä määrin niiden fenotyyppi todella muistuttaa asukas makrofagien vaikuttaa enteerisellä neuronien, erityisesti reseptorin ilmentymistä makrofageissa voi olla ylös- tai alaspäin säännellään ne on eristetty luonnollisesta ympäristöstään. Siksi päätimme kehittää tekniikkaa, joka antaisi meille mahdollisuuden tutkia vaikutusta nikotiinin aktivoinnista asuvan makrofagien niiden luonnollisessa ympäristössä. Kuten makrofagit aktivoituvat ATP, tunnettu vaara signaali immuunisolujen [8,9], paljastaa kasvu solunsisäisen Ca 2 + elävät Ca 2+ kuvantaminen valittiin tutkimaan asuva makrofagien koskemattomassa tasainen arkki hiiren vatsa valmisteita. Tämä antoi meille mahdollisuuden vertailla ATP herätti Ca 2+ transientit ([Ca 2 +] i) makrofageissa sijaitsee sileän lihaksen kerrokset ennen käyttöä ja sen jälkeen nikotiinia. Olemme lisäksi käytetty useita antagonisteja, joilla tiedetään mieltymyksiä tiettyihin nAChR alayksiköiden jotta edelleen mekanistinen oivalluksia roolit nikotiinin reseptoria ilmentävien makrofagit varten CAIP suolistossa. Nämä farmakologiset havainnot vahvistettiin immunohistokemiallisesti. Lopuksi analysoidaan osuutta asuva makrofagien, jotka ovat välittömässä läheisyydessä, kolinergisten hermosäikeitä. Ethics lausunto Kaikki hiiri toimivat tehtiin Saksan suuntaviivojen eläinten hoito ja hyvinvointi (Deutsches Tierschutzgesetz) ja hyväksytty Baijerin eettisen komitean (Regierung Oberbayern, joka toimii laitos- ja Käytä komitean Technische Universität München) mukaan 4 § ja 11 § Deutsches Tierschutzgesetz viitenumerolla 32 -568-2. kudosnäytteitä Male 12-16viikko ikäisiin C57BL /6-hiiriä (Charles River, Sulzfeld, Saksa) tapettiin katkaisemalla kaula. Vatsa kerättiin jääkylmään Krebs, joka sisälsi (mM) 117 NaCI, 4,7 KCI, 1,2 MgCI 2 6 H 2O, 1,2 NaH 2PO 4, 25 NaHCO 3, 2,5 CaCl 2 2 H 2O ja 11 glukoosia ja säädettiin pH-arvoon 7,4. Vatsa avattiin suurempaa kaarta pitkin ja pestiin jääkylmällä Krebsin. Alle mikroskooppinen tarkastus, vatsa oli kiinnitetty alas sylgard lautasen ja limakalvoa ja submukoosan poistettiin varovasti. Aikana leikkely, kudos jatkuvasti perfusoitiin jääkylmällä Krebsin ratkaisu, jolla varmistetaan elinkelpoisuuden kudosta. Vain etummaisessa tai lantio puolet vatsa oli kiinnitetty pienellä sylgard rengas, jonka keskellä on aukko 100 x 200 mm 2. Flat levyt hiiren vatsa valmistelut olivat inkuboidaan modifioitu Krebs liuokseen (117 NaCl, 4,7 KCI, 1,2 MgCI 2 6 H 2O, 1,2 NaH 2PO 4, 20 NaHCO 3, 2,5 CaCl 2 2 H 2O ja 11 glukoosia), joka sisälsi 30 pM fluoresoivaa kalsiumin indikaattorina Fluo 4-asetoksimetyyli (AM) (Invitrogen), ja 1,25 mM probenesidi (Sigma-Aldrich, Schnelldorf, Saksa) 2 tuntia huoneen lämpötilassa pimeässä ja kaasutettu karbogeenikaasu (95% O 2-5% CO 2). Sylgard rengas asennettiin tallennus kammiossa herakalvon puolella vatsan pohjaan päin kammion. Kammio oli kytketty perfuusion järjestelmä mahdollistaa jatkuvan perfuusion karbogeenikaasu kaasut Krebs liuokseen huoneenlämpötilassa. Huuhtelujakso 1,5 h annettiin ennen kokeen aloittamista. Kudos kammio kiinnitettiin käänteisen epifluoresenssimikroskooppia (Zeiss Axio Observer A1, Carl Zeiss, Jena, Saksa) varustettu nopea yksivärinen kamera (Zeiss AxioCam HSM) ja ohjelmiston (Zeiss Axio Vision 4.8) hankintaan ja analysointiin. Fluo-4 viritettiin käyttämällä sinistä valoa lähettävä diodi (LED) Luxeon III (3W, 470 nm hallitseva aallonpituus, Philips Lumiled, Phillips, Hambur, Saksa) ja Fluo-4 signaalit havaittiin suodattimella kuution F26-514 Kirkas Line FITC BP (eksitaatio: HC475 /35, Dichroic: 499, päästöt: HC530 /43, AHF ANALYSENTECHNIK, Tübingen, Saksa) käyttäen X20 tavoite (A-suunnitelma, NA = 0,25, Zeiss). Järjestelmä mitattiin suhteelliset muutokset fluoresenssissa (Δ F /F) Fluo-4 muutosten seuraamiseksi solunsisäisen kalsiumin ([Ca 2 +] i). Ca 2+ häiriö- kirjattiin alkaen 3 s ennen paikallista antoa ATP yhteensä 14,5 s kuvanopeus 2 Hz ja valotusaikaa 200 ms. käytetään ATP välineenä makrofagiaktivaation, koska se on merkki vaarasta vapautuu paikallisesti tulehduskohtaan [8,9], ja koska ATP tiedetään aiheuttavan sytokiinin eritystä makrofageista kautta lisäystä [Ca 2 +] i [10-12]. ATP (Sigma-Aldrich) ja nikotiini (Sigma-Aldrich) on paikallisesti soveltaa paine paiskautuu kahdesta mikropipetit joiden pituus on 200 ms ja 10 s, vastaavasti. Asento mikropipetit varmistettiin, että ulos volyymit kattaa samat kudoksen alueille. Mikropipetit täytettiin 1 mM ATP ja 100 uM tai 10 uM nikotiini liuotetaan Krebsin liuokseen, joka sisältää 1,25 mM probenesidiä. Mukaan aiemmin julkaistu kalibrointikäyrien perusteella arvioimme, että mitään ainetta levitetään paineen poisto pulsseja laimennetaan noin 1:10, kun se saavuttaa kudoksen [13]. Paikallinen anto ATP ja nikotiinin sallitaan mittaaminen vastausten useilla alueilla (tyypillisesti 4-5) samassa kudoksessa. Asema alueiden kudos dokumentoidaan koordinaatiston näkyy mobiili mikroskoopin vaiheessa. Vaikutus nikotiinin ATP aiheuttama kalsiumin häiriö- jälleen tutkittiin samassa alueilla jälkeen lisäämällä erilaisia nAChR antagonisteja. Tallennuksen jälkeen vastaukset, makrofagit visualisoitiin tärkeää inkuboimalla kudos allofykosyaniini (APC) leimattua rotan anti-hiiri anti-F4 /80-vasta-ainetta (1: 250, eBioscience, Frankfurt, Saksa) 1 tunnin ajan, huoneenlämpötilassa tumma ja kaasutettiin karbogeenikaasu. Kudos pestiin 15 minuutin ajan. Mikroskooppi vaihe sijoitetaan uudelleen löytää alueita, joilla me tallennettu. Kuvia leimattuja makrofagien hankittiin käyttämällä punaista Z-LED P4 (3,5 W) magnetointilähdettä (625 nm hallitseva aallonpituus, Seoul Semiconductor) ja suodatin kuution F46-006 ET suodatin setti (eksitaatio: ET 620/60, Dichroic: 660, päästöt: ET700 /75, AHF ANALYSENTECHNIK, Tübingen, Saksa). Peittokuvaa, Fluo-4 signaalit ja kuvat F4 /80 positiivinen makrofagien pystyimme analysoida vastauksia yksittäisiin makrofageissa. etiketti kudoksen asukas makrofagien, ensin käytetty elintärkeitä merkinnöissä protokollaa edellä kuvatulla tavalla. Kudos kiinnitettiin sitten yön yli huoneen lämpötilassa liuokseen, joka sisälsi 4% formaldehydiä ja 0,2% pikriinihappoa 0,1 M fosfaattipuskurissa, pestiin 3 x 10 minuuttia fosfaattipuskurissa ja lopuksi inkuboitiin 1 tunnin ajan liuoksessa, joka sisälsi 0,5% Triton X- 100, 0,1% NaN 3, 4% hevosen seerumissa PBS: ään liuotettua (kaikki Sigma-Aldrich). Merkitä kolinergisen varicosities, kudosta inkuboitiin yön yli esto, joka sisälsi vuo- hen anti-vesicular asetyylikoliinin kuljettajan (VAChT; 1: 1000, Merck-Millipore, Darmstadt, Saksa). Kudokset pestiin 3 x 10 min PBS: ssä ja inkuboitiin 1,5-2 h eston, joka sisälsi Cy3-leimattua anti-vuohi-vasta-aine (1: 500, Dianova, Hampuri, Saksa). Kudokset pestiin 3 x 10 minuuttia PBS: ssä ja asennetaan antihäipyminen aine (20% PBS /NaN 3, 80% glyseroli) poly-L-lysiinillä päällystettyihin dioja ja peitetään katseluun. merkitä β2 nAChR kudoksessa asuva makrofageissa, limakalvon vapaa mahalaukun koko vuori valmisteiden villityypin ja p2 nAChR knock-out [14] hiiriä kiinnitettiin 10 min jääkylmään PBS-liuosta, joka sisälsi 4% paraformaldehydiä (PFA) . Kudokset pestiin sitten 2 x 10 min PBS: ssä ja inkuboitiin 2 h PBS: ssä, joka sisälsi 1% proteaasia Albumin Bovine Fraction V (BSA; Serva, Heidelberg, Saksa) ja 10% normaalia aasi seerumia (NDS, Jackson Immuno- Research, Pennsylvania, USA). Kudoksia inkuboitiin 36 h PBS: llä, joka sisälsi 1% BSA: ta, 5% NDS, rotan anti-hiiri-F4 /80 (1: 500; klooni BM8, BioLegend, San Diego, USA) ja kanin anti-hiiri-β2 nAChR (1: 200; Santa Cruz Biotechnology, Heidelberg, Saksa). Kudokset pestiin 3 x 10 min PBS: ssä ja inkuboitiin 1 h PBS: ssä, joka sisälsi 1% BSA: ta, 5% NDS, Alexa Fluor® 647-leimattua vuohen anti-rotta-vasta-ainetta (1: 1000, Jackson Immuno- Research, Pennsylvania, USA) ja Cy3-leimattua aasin anti-kani-vasta-ainetta (1: 1000; Chemicon, Millipore, Billerica, USA). Kudokset pestiin 3 x 10 minuuttia PBS: ssä ja asennetaan hitaasti fade reagenssia (Invitrogen Life Technologies, Gent, Belgia) poly-L-lysiinillä päällystettyihin dioja ja päällystettiin varten viewing.To etiketin α7nAChR kudoksessa asuva makrofageissa, eli pala hiiren mahan ja sykkyräsuoli alistettiin tärkeää merkintöjä käyttäen Cy5-leimattua α-Bungarotoksiini (Invitrogen) on 0,1 ug /ml RPMI 1640 -elatusaineessa (Lonza, Basel, Sveitsi) 4 ° C: ssa 15 min [4]. Kudokset kiinnitettiin sitten PBS: ssä, joka sisälsi 4% PFA. Limakalvon ja submukoosassa poistettiin ja kudoksia inkuboitiin 2 h esto, joka sisälsi 1% BSA: ta. Kudoksia inkuboitiin sitten 60 h esto, joka sisälsi rotan anti-hiiren F4 /80 (klooni BM8, BioLegend), jota seurasi 3 x 10 min PBS-pesun ja inkuboitiin 1 h PBS: ssä, joka sisälsi 1% BSA: ta ja Cy3-leimattua anti-rotta-vasta-ainetta (1: 1000, Jackson Immuno-, Pennsylvania, USA). Lopuksi kudokset pestiin 3 x 10 minuuttia PBS: ssä ja asennetaan hitaasti fade aineen poly-L-lysiinillä päällystettyihin dioja ja peitetään katseluun. kuvat otettiin käyttäen LSM 510 (Carl Zeiss) -konfokaalimikroskoopilla Plan-Neofluar x40 /1.3 Öljyn DIC ja Plan Apochromat x63 /1,4 Oil DIC tavoitteita. Laser aallonpituuksilla 543 nm ja 633 nm: ssä käytettiin magnetointi fluoroforien Cy3 ja APC tai Cy5, vastaavasti. Cy3 ja APC tai Cy5 signaaleja havaittiin käyttäen suodatinta asettaa BP 565-615 IR ja BP 650-710 IR, vastaavasti. Kuva pinot varten kvantitatiivisen analyysin avulla x63 tavoite skannattiin XY 1024 × 1024 että pinta-ala on 95,5 x 95,5 um 2. Ensimmäinen ja viimeinen optisten viipaleita otettiin yläosassa ja alaosassa ulkopinnan makrofagi. Optinen syvyys kunkin viipaleen oli 900 nm. Kaksi peräkkäistä viipaleita limittäin 500 nm. Yleensä välillä 9 ja 16 viipaletta syntyi tuloksena skannauksen syvyys 3,2-6,0 mikrometriä, joka sisältää välillä 1-3 makrofagien ja VAChT säikeet ylitys makrofagit. Kuvapinot analysoitiin Image J Pro. Kuvia β2 ja α7 nAChR-leimattua makrofagien otettiin käyttäen x63 tavoite ja skannattu XY 1024 × 1024 että pinta-ala on 95,5 x 95,5 um 2. Optinen syvyys kuvien oli 900 nm. Voit estää aktiopotentiaalin eteneminen neuronien tetrodotoksiiniin (Biotrend, Köln, Saksa) lisättiin perfuusio Krebsin liuosta 1 uM. Farmakologisen karakterisoinnin, seuraavat nikotiini- salpaajia lisättiin Krebsin liuokseen perfuusio kudos: 200 uM heksametoniumkloridin (Sigma-Aldrich), 100 uM mekamyyliamiinin (Sigma-Aldrich), 10 uM dihydro-β-erytroidiini (DHBE; Sigma-Aldrich) , 100 nM, 3 uM ja 10 uM α-Bungarotoksiini (ABGT, Tocris) ja 10 nM ja 100 nM methyllycaconitine (MLA, Tocris). Heksametonium, mekamyyliamiinin ja DHBE testattiin pitoisuuksina, jotka pystyivät poistamaan nikotiini- nopeasti kiihottavien postsynaptisiin potentiaalit marsun myenteeri- neuronien [15] kantavassa käyttö 10 nM ja 100 nM MLA perustuu pitoisuus vastaavasti käyttää estämään α7 alayksikön sisältävien nAChR [16] ja käyttää estämään IL-6 eritys eristetty peritoneaalimakrofageille [3]. Tietojen analysointi ja tilastot maksimaalinen suhteelliset muutokset fluoresenssissa (Δ F /F) vastauksena ATP hallinto ilmaistiin%: n nousu yli perustason fluoresenssi ennen ATP antoa. Tilastolliset analyysit tehtiin Sigma Plot 9,0 (Systat Software Inc, Erkrath, Saksa). Tiedot esitetään viiksi käyriin mediaani ja 25 th ja 75 th persentiilit sekä 10 th ja 90 th persentiilit. Ei normaalisti jakautunut tulospareille analysoitiin Wilcoxonin testi. Erot katsottiin tilastollisesti merkitsevä P tulokset Spatial suhde kudoksen asukas makrofagien ja kolinerginen varicosities hiiren vatsassa käyttää konfokaalimikroskopia arvioida läheisyydessä välillä F4 /80-positiivisia makrofageja ja VAChT-positiivisten suonikohjuja kolinergisten hermosyiden hiiren mahaan (kuvio 1A). Yksityiskohtainen analyysi paljasti, että 83% 41 makrofagien tutkittu sijaitsevat 900 nm ainakin yhden suonikohjuja kolinergisten hermojen kuidun (kuvio 1 B-C). Microejection ATP indusoi voimakkaan, alkaa nopeasti [Ca 2 +] i ohimenevä makrofageissa, joka saavutti huippunsa 8-10 sekunnin kuluttua sovellus seuraa hidas palaaminen perustasolle [Ca 2 +] i tasoja (kuvio 2A ja Movie S1). Koska ei ollut aina täydellinen toipuminen lähtötasolle aikana tallennusaika, maksimaalinen [Ca 2 +] i-signaalin käytettiin analyysiin kaikissa kokeissa. Ei takyfylaksi havaittu, koska maksimaalinen amplitudi [Ca 2 +] i signaalin vastauksena toiseen ATP hallinto, 10 minuuttia ensimmäisen jälkeen, ei poikennut alkuperäisestä maksimaalinen vaste (kuvio 2B). vaikutus nikotiinin [Ca 2 +] i signaaleja kudoksessa asuva makrofageissa vaikutuksen tutkimiseksi nikotiinin ATP herätti [Ca 2+] i merkeistä microejected nikotiinia 10 sekunnin ajan ja välittömästi uudelleen käyttöön ATP päälle samalla alueella. Yksi järkevä käyttää nikotiinia selektiivinen, ei-hajoavat nAChR-agonistin oli jäljitellä nikotiinireseptorin aktivointi asetyylikoliinin vapautumista kolinergisten neuronien. Analysoidaan muutokset [Ca 2 +] i kaikissa makrofageissa paljasti, että nikotiini vähensi ATP herätti [Ca 2 +] i signaalit (kuvio 2D ja F). Tarkempi analyysi vaikutuksista nikotiinin ATP aiheuttama [Ca 2 +] i häiriö- paljasti kolmen populaation makrofagien (kuva 2D-G). Nikotiini 10 ja 100 uM heikennettyjä ATP-herätti [Ca 2 +] i signaalit 55% ja 65% makrofagien, vastaavasti. Esiin tulee [Ca 2 +] i-signaalin pysyi ennallaan 36% ja 28% makrofagien soveltamisen jälkeen 10 uM ja 100 uM nikotiini, vastaavasti. Vuonna pieni joukko, 10 uM ja 100 uM nikotiini voimisti ATP-herätti [Ca 2 +] i-vaste (9% ja 7% makrofagien, vastaavasti). välttämiseksi bias lisäksi analyysi perustuu nikotiinin vaikutukset kaikkiin makrofageissa riippumatta siitä, onko ATP aiheuttama [Ca 2 +] i signaali vähentynyt, lisääntynyt tai ennallaan. rooli neuronien nikotiini- herätti vaimennus makrofagiaktivaatiota Nikotiini aktivoi suoraan enteerisen neuronien ja olemme käsitelleet mahdollisuutta, että aktivointi lähellä olevaan myenteeri- neuronien vaikutti heikennetty [Ca 2 +] i vasteita makrofageissa. Emme löytäneet todisteita tällaista epäsuoraa estävää koulutusjakson koska vaimennus ATP-herätti [Ca 2 +] i signaalin nikotiinin pysyi läsnäollessa tetrodotoksiiniin (kuvio 2C). On huomionarvoista kuitenkin, että osa niistä makrofagien jossa nikotiini ei muuttanut ATP herätti [Ca 2 +] i signaalit vähentynyt voimakkaasti (28% vs. 6%). Samaan aikaan, makrofagit jossa nikotiinin esti tai voimistua ATP herätti [Ca 2 +] i signaaleja nousi 65%: sta 71% ja 7%: sta 23%: iin. makrofagien aktivoituminen ATP ja inhibitio ATP vasteen 100 uM nikotiinin luotettavasti kirjattiin kussakin 21 valmisteiden kuviossa 2F. Tämä antoi meille mahdollisuuden tehdä antagonistien tutkimukset ilman tarvetta restudy inhiboivan vasteen, jotka makrofagit käsitelty antagonisteja. Lisäksi olemme heikennä määrää tallennuksen kausien tasolle, joka ei vaaranna signaalin voimakkuuden ja taattu toistettavissa ATP vastauksia. Jotta voidaan tutkia nAChR-alayksiköt osallistuvat estävä vaikutus nikotiinin, testasimme viisi erilaista salpaajia, joilla on tunnettu alayksikön mieltymykset [17] (kuvio 3). Estovaikutus nikotiinin ATP-herätti [Ca 2 +] i vasteita pysyi muuttumattomana, kun läsnä on ei-selektiivinen ganglioninen nAChR antagonisti heksametoniumkloridin The α3β4 nAChR-mieluummin nisti mekamyyliamiinia tai α7 nAChR-parempana antagonistit α-Bungarotoksiini ja methyllycaconitine (kuvio 3A). Kuitenkin β2 nAChR-suosiminen antagonisti di-hydro-β-eryhtroidine päinvastaiseksi estävää vaikutusta nikotiinin ATP-herätti [Ca 2 +] i vasteita makrofageissa (kuvio 3B). Vaikka käytimme ABGT pitoisuuksina, jotka on kuvattu luotettavasti estää α7 nAChR neuronien ja eristyksissä keuhkorakkuloiden makrofagien [18], olimme huolissaan siitä, että se ei ehkä pysty antagonisoida estävää vaikutusta nikotiinin vuoksi epäsuotuisa kilpailun on sitoutumiskohta. Näin näyttää kuitenkin epätodennäköiseltä, koska jo pitoisuuksilla 3 uM ja 10 uM ABGT ei kyennyt kääntämään estävää vaikutusta nikotiinin ATP herätti [Ca 2 +] i vasteita (kuvio 3A). ABGT ei myöskään kääntää vaimennus herättämän 10pM nikotiini (kuvio 3C). leimaaminen β2 muttei a7-nAChR kudoksessa asuva makrofageissa Tuottaa lisätodisteita osallistumista β2 nAChR mutta ei α7 nAChR vuonna estävää vaikutusta nikotiinin ATP-herätti [Ca 2 +] i vastauksia, immunohistokemiallinen merkitseminen p2 nAChR ja elintärkeä merkintöjä α7 nAChR mukaan ABGT kotimaisissa makrofageissa mahan muscularis tehtiin (kuva 4). Suurin osa asukas makrofagien mahalaukun muscularis oli p2 nAChR-immunoreaktiivinen (kuvio 4A), joka tukee havaittu antagonistinen vaikutus DHBE nikotiinireseptoreihin inhibitio ATP vastauksia. Käytetty vasta-aine p2 nAChR on erityinen, koska puuttuminen p2 nAChR immunoreaktiivisuus on β2 nAChR hiiressä (kuvio 4B). Vital merkintöjä α7 nAChR mukaan ABGT paljasti puuttuminen α7 nAChR kudokseen asuva makrofagien hiiren mahaan (kuvio 4C), sen sijaan, suoliston makrofagit leimattiin ABGT (kuvio 4D). Puute α7 nAChR vatsassa muscularis asukas makrofagien on vahvistettu, ettei antagonismi α7 nAChR-mieluummin salpaajat ABGT ja MLA sen estävää vaikutusta nikotiinin ATP herätti [Ca 2 +] i vasteita näissä soluissa. keskustelu Toistaiseksi vaikutus nikotiinin on tutkittu vasta eristetyissä makrofageissa tai makrofaagisolukantoja. Tässä olemme kehittäneet in vitro kriteerinä käytetään määrittelemään lähellä välillä makrofagien ja kolinergisten hermosyiden (900 nm etäisyys) yhtyy käsitteeseen extrasynaptic viestintä . Tämän käsitteen diffuusioon perustuva tilavuus lähetys voi tapahtua 100 nm um etäisyydet lähde ja kohde [19]. Oletetaan, että useimmat, elleivät kaikki, kolinergisten hermojen läheisyydessä makrofageihin alkunsa myenteeri- hermosolujen elinten pohjautuvat aiempien havainto, että vagaalisia kuidut eivät ota suoliston asukas makrofageissa [3] Tämä on myös tukevat havainnot, että mahalaukun vagaalisia efferent kuidut lähes yksinomaan päättyvät enteerinen hermosolun [20], jossa ne aktivoivat enemmistön myenteeri- neuronien kautta nikotiinireseptoreihin [15]. CAIP edustaa fysiologista valvontajärjestelmän makrofagiaktivaatio aikana tulehdustiloja. Tulehdusta estävä vaikutus CAIP aktivaation on osoitettu in vivo Olemme kuitenkin valita melko konservatiivinen tulkinnan tietomme ja päätellä, että β2 nAChR kriittisesti mukana nikotiini estämällä makrofagien toimintaa, vaikka on todennäköistä, että käytetyn pitoisuuden tutkimuksessamme DHBE mieluiten lohkot α4β2 nAChR. Meidän valmiste soveltuu erinomaisesti käsitellä tulevissa tutkimuksissa mahdolliseen osallistumiseen α7β2 nAChR tutkimalla vaikutus nikotiinin kudoksessa asuva makrofageihin α7 nAChR, p2 nAChR ja α7β2 nAChR kaksinkertainen knock-out hiirillä. Samanlaisia strategioita olisi käytettävä tutkia merkittävyyden α4β2 nAChR. On tärkeää huomata, että antagonistinen toiminta DHBE ei ole selektiivinen makrofagien kuin DHBE, kuten heksametoniumkloridin ja mekamyyliamiinia estää myös nikotiini- synapsien mahalaukun myenteeri- neuronien [15]. Kuitenkin nikotiinireseptoreihin makrofageissa on oltava erilaisia ominaisuuksia kuin enteerinen neuronien koska kumpikaan heksametoniumkloridin eikä mekamyyliamiinia päinvastaiseksi nikotiinin aiheuttama vaimennus ATP-mieleen vasteita makrofageissa. Heksametonium ja mekamyyliamiinin vaikutus perustuu niiden toimista tukkimalla pore nikotiinihapon kanavan [23-25]. Niiden kyvyttömyys kääntää nikotiinin inhibitio makrofagien voi ehdottaa, että on olemassa epätyypillinen nAChR makrofageissa. Todellakin, kirjaaminen Ca 2+ transientteja vastauksena nikotiinin annon paljasti kasvanut [Ca 2 +] i vain 13% makrofageja (6 out of 45 makrofageja; tuloksia ei ole esitetty). Tämä populaatio on paljon pienempi, että osuus makrofagien jossa nikotiini moduloitu ATP herätti [Ca 2 +] i. Runsas todistusaineisto viittaa siihen, että α7 nAChR keskeinen rooli nikotiinin aiheuttama väheneminen makrofaagien sytokiinituotannossa [2,3,6,26]. Aiemmin olemme todellakin osoittaneet, että nikotiini ei vähentää TNF-α tuotannon vatsakalvon makrofageissa α7 nAChR hiirten [6], kun taas anti-inflammatorisen vaikutuksen ohutsuolessa on Kiertäjähermo stimulaation mallissamme postoperatiivisten ileus menetetään näissä KO hiiret [4]. Esillä olevassa tutkimuksessa kuitenkin vaikutus nikotiinin ATP aiheuttaman makrofagiaktivaatio ei estänyt α7 nAChR mieluummin salpaajat ABGT ja MLA, vastaan puhuu osallistumista α7 nAChr. Tätä tukee lisäksi merkkauksen puuttuminen mahalaukun muscularis makrofagien α7 nAChR-suosiminen α-Bungarotoksiini. Suolessa, mutta teimme tarkkailla α-Bungarotoksiini positiivinen leimattu muscularis makrofagien [4 ja tämän tutkimuksen], mikä osoittaa, aluekohtaisia eroja fenotyypin näiden immuunisolujen. Vaikka ilmeinen ABGT herkkien α7 nAChR tutkimuksessamme näyttää ristiriitaiselta aikaisempien havaintojemme, olemme melko varmoja, että nämä tiedot eivät johdu kyvyttömyydestä ABGT kilpailla sitoutumiskohtaa koska sama pitoisuus käytetty meidän opintokokonaisuudet hermosolujen α7 nAChR aivoissa ja eristetty keuhkorakkuloiden makrofageissa [18].
Tulokset
Johtopäätös
situ
osoitus eston makrofagiaktivaatiota nikotiinin viittaa toiminnallisen välinen signalointi kolinergisten neuronien ja makrofagit vatsassa. Tiedot viittaavat siihen, että β2 alayksikköä nAChR on kriittisesti mukana nikotiinin aiheuttaman eston näiden asukas makrofagien.
Johdanto
Methods
Kalsium kuvantamisen
immunohistokemia
Konfokaalimikroskopia ja kuva-analyysi
farmakologian
< 0,05. N
numerot suluissa viittaavat numerot makrofagien /kudosten tutkittu, eli tulos perustuu tallenteita 20 makrofagien 5 kudoksissa (yhtä kuin 5 eläintä) esitetään (20/5).
toistettavuus ATP mieleen [Ca 2 +] i signaaleja kudoksessa asuva makrofageissa
farmakologinen karakterisointi inhiboiva vaikutus nikotiinin kudoksen asuvat makrofagien
mallissa hiiren vatsan lihaksia myenteeri- plexus valmiste tutkia vaikutusta nikotiinin kudoksen asuville makrofagien niiden luonnollisessa ympäristössä. Tämä esillä olevassa tutkimuksessa on siis ensin osoittaa, että nikotiini suoraan estää aktivoitumisen kudoksen asuvan makrofagien kautta β2 alayksikön sisältävien nAChR ja siten luo perustan toiminnallisen signalointi kolinergisten neuronien ja makrofagit suolistossa. Meidän johtopäätöstä tukee useita rivejä todisteita. Ensinnäkin ATP herätti [Ca 2 +] i transienteilta makrofageissa alentaa merkittävästi nikotiinin vaikka läsnä hermo esto tetrodotoksiiniin. Toiseksi nikotiinin aiheuttama vaimennus ATP vastausten makrofageissa purettiin DHBE, mutta ei heksametoniumilla, mekamyyliamiinia, ABGT tai MLA. Farmakologiset havainnot tukivat osoitus β2-positiv mutta ABGT-negatiivinen nAChR alayksiköiden mahalaukun makrofageissa. Kolmanneksi, suurin osa makrofagit olivat suorassa läheisyydessä suonikohjut kolinergisten hermosyiden. Samanlaisia lähellä makrofagien hermosyiden havaittiin rotan suoliston muscularis [3].
by vagushermostimulaatiolle hiirimalleissa sepsiksen ja leikkauksen jälkeinen suolentukkeuma [2,3] ja in vitro
nikotiinin annon eristyksissä , lipopolysakkaridi stimuloduista makrofageissa [1-4,21]. Tässä tutkimuksessa havaitsimme, että nikotiini vähentää ATP-indusoidun solunsisäisen Ca 2+ asuvat makrofageissa vatsassa. Kiinnostavaa kyllä, tämä vaikutus oli päinvastainen jonka β2 nAChR alayksikköä mieluummin antagonisti DHBE viittaa osallistumista tämän alayksikön nikotiini välittämää modulaatioon asukas makrofageissa. Tämä havainto on johdonmukainen aiempaan toteamukseen DHBE käänteinen nikotiinin aiheuttaman eston tuumorinekroositekijä-α (TNF-α) vapautumista ja lisääntynyt fagosytoosin eristetyissä peritoneaalimakrofageille [6]. Linjassa, sytokiinien tuotantoa ihmisen neuroblastoomasolulinja stabiilisti transfektoitu α4β2 nAChR merkittävästi vähentää nikotiinin esikäsittely [22]. Vaikka nämä tiedot viittaavat siihen, että α4β2 nAChr saattaa välittää vaikutus nikotiinin ATP aiheuttama solunsisäisen Ca 2+, viimeaikaiset todisteet osoittavat, että β2 alayksiköt voivat myös koota muiden α alayksiköiden, kuten α7 alayksiköt. Tuoreessa julkaisu keskustella elektrofysiologisiin ominaisuuksia α7β2 nAChR ilmaistuna ihmisen epiteelisolujen solulinjoissa, osoittivat, että pieninä pitoisuuksina DHBE antagonisoivat α7β2 nAChR mutta ei a7-nAChR [16]. Nämä ja muut tiedot farmakologinen profiili DHBE ehdottaa, että DHBE on erittäin selektiivinen p2 nAChR mutta ei luotettavasti erottaa eri kokoonpanot α3, α4 tai α7 kanssa β2. Kuitenkin meidän immunohistokemiallinen ja elintärkeä merkintöjä mahalaukun asuva makrofagien eivät puolla osallistumista α7 nAChR koska mahalaukun makrofagit eivät leimattiin ABGT mutta ilmaisi β2 nAChR alayksikköä.
 Nailed It:Highlights from the Gut Stuff
Nailed It:Highlights from the Gut Stuff
 Sjögrenin oireyhtymä:mistä se johtuu ja miten siitä voi selvitä luonnollisesti
Sjögrenin oireyhtymä:mistä se johtuu ja miten siitä voi selvitä luonnollisesti
 400 nyt sairastuneena vatsataudin puhkeamiseen
400 nyt sairastuneena vatsataudin puhkeamiseen
 5 yksinkertaista tapaa voittaa happorefluksi tänään
5 yksinkertaista tapaa voittaa happorefluksi tänään
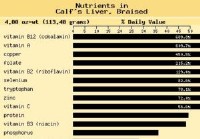 SCD-resepti:Sitruunapippuri-maksapihvi
SCD-resepti:Sitruunapippuri-maksapihvi
 Maaliskuu on kansallinen kolorektaalisyöpätietoisuuskuukausi
Maaliskuu on kansallinen kolorektaalisyöpätietoisuuskuukausi
 Yhdeksän ruoansulatussairauksien myyttiä
Lääketieteelliset arvioijat ja toimittajat:Jay W. Marks, MD ja Melissa Conrad Stöppler, MD Tutkijat ovat vasta hiljattain alkaneet ymmärtää monia, usein monimutkaisia, ruoansulatusjärjestelmään vaiku
Yhdeksän ruoansulatussairauksien myyttiä
Lääketieteelliset arvioijat ja toimittajat:Jay W. Marks, MD ja Melissa Conrad Stöppler, MD Tutkijat ovat vasta hiljattain alkaneet ymmärtää monia, usein monimutkaisia, ruoansulatusjärjestelmään vaiku
 Vatsavaivoja, jotka eivät liity autismiin, tutkimustulokset
Viimeisimmät neurologiauutiset Lisää todisteita, jotka yhdistävät monon multippeliskleroosiin Pienikin liikunta voi hidastaa Parkinsonin tautia Lääke saattaa auttaa helpottamaan autismin aistillisia
Vatsavaivoja, jotka eivät liity autismiin, tutkimustulokset
Viimeisimmät neurologiauutiset Lisää todisteita, jotka yhdistävät monon multippeliskleroosiin Pienikin liikunta voi hidastaa Parkinsonin tautia Lääke saattaa auttaa helpottamaan autismin aistillisia
 Mikä Weber-Christian Disease on?
Weber-Christian tauti on harvinainen tulehduksellinen sairaus, joka vaikuttaa kehon rasvakudoksiin. Häiriö ilmenee iholla punaisina tai purppuranpunaisina arkina, kohonneena kyhmynä yleensä reisissä j
Mikä Weber-Christian Disease on?
Weber-Christian tauti on harvinainen tulehduksellinen sairaus, joka vaikuttaa kehon rasvakudoksiin. Häiriö ilmenee iholla punaisina tai purppuranpunaisina arkina, kohonneena kyhmynä yleensä reisissä j