Maag inhoudsanalyse (SCA) en meer recent stabiele isotopen analyse (SIA) geïntegreerd met isotopen mengen modellen hebben gemeenschappelijke methoden voor de voeding studies worden en geven inzicht in de foerageerecologie van zeevogels. Beide methoden hebben nadelen en vertekeningen die kunnen leiden tot moeilijkheden bij het kwantificeren interjaarlijkse en soort-specifieke verschillen in voeding. We gebruikten deze twee methoden om gelijktijdig te kwantificeren de-chick opfokvoer van Chinstrap ( Pygoscelis antarctica Visum:. Polito MJ, Trivelpiece WZ, Karnovsky NJ, Ng E, Patterson WP, Emslie SD (2011) Integratie van de Maag inhoud en stabiele isotopen analyses om de Diëten van Pygoscelid Penguins kwantificeren. PLoS ONE 6 (10): e26642. doi: 10.1371 /journal.pone.0026642 Editor: André Chiaradia, Phillip Island Nature Parks, Australië Ontvangen: 2 juni 2011; Aanvaard: 30 september 2011; Gepubliceerd: 28 oktober 2011 Dit is een open-access artikel, vrij van auteursrechten, en mag vrij worden gereproduceerd, gedistribueerd, verzonden, gewijzigd, gebouwd op, of anderszins gebruikt door iedereen voor elk rechtmatig doel. Het werk wordt onder de Creative Commons CC0 publieke domein toewijding ter beschikking gesteld Financiering:. Dit onderzoek werd gefinancierd door de Amerikaanse AMLR Program (http://swfsc.noaa.gov) en de Amerikaanse National Science Foundation Bureau van Polar Programs (www.nsf.gov) subsidies aan S. Emslie en W. Trivelpiece. De financiers hadden geen rol in de studie design, het verzamelen van gegevens en analyse, besluit te publiceren, of de voorbereiding van het manuscript Competing belangen:.. De auteurs hebben verklaard dat er geen tegenstrijdige belangen bestaan Introductie Maag inhoudsanalyse (SCA) is een van de meest voorkomende methoden voor de voeding analyse en geeft inzicht in de foerageerecologie van zeevogels en de verspreiding, omvang en demografie van hun prooi [1], [2]. Vroege studies vaak betrokken offeren dieren maaginhoud [3] onderzocht, terwijl nog een niet-destructief, maar invasief, "lavage" techniek om regurgitatie, die meestal wordt toegepast [4], [5]. Wanneer teruggewonnen maaginhoud relatief onverteerd, is het mogelijk om de samenstelling en de frequentie van voorkomen prooisoorten schatten en vaak meten, wegen en geslacht individuele prooi [3]. Bovendien, het identificeren en meten van harde prooien resten, zoals inktvis snavels en otolieten, kan informatie over de grootte en massa van prooidieren verstrekken wanneer prooi heeft geheel of gedeeltelijk verteerd [6], [7], [8]. Er zijn inherente nadelen en vooroordelen bij het gebruik van SCA om zeevogel diëten te kwantificeren. Deze techniek is het meest gebruikt tijdens kuiken opfok bij volwassenen brengen voedsel aan wal om hun kuikens; dus minder bekend is over de voeding van zeevogels buiten het broedseizoen [2]. maaginhoud ook wijzen op een 'snapshot' van de afgelopen dieet van een individu (8-16 uur) en kan zeer variabel zijn, die grote steekproefomvang statistische verschillen tussen soorten, regio's en /of tijd te onderzoeken [3], [9], [ ,,,0],10]. Bovendien wordt voorgespannen naar SCA Language voeding items en buit niet gemakkelijk doet verteren, zoals zooplankton en kan de hoeveelheid zacht lichaam prooi, zoals vis en inktvis [11], [12] onderschatten. Hoewel harde prooi blijft van maaginhoud of pellets informatie verstrekken over prooidieren samenstelling van deze gegevens zijn vaak moeilijk te integreren in de totale voeding samenstelling schattingen [6], [8], [13]. Recente ontwikkelingen in de stabiele isotoop analyse (SIA) en isotoop mengen modellen hebben laten zien een grote belofte in het kwantificeren van de voedingskundige samenstelling van zeevogels [14], [15]. Isotopische analyses zijn gebaseerd op het concept dat dieren "zijn wat zij eten" met weefsel stabiel stikstof (δ 15N) en koolstof (δ 13C) verhoudingen weerspiegelen dieet bij de synthese [16]. Bijvoorbeeld, veren zijn metabolisch inert na de synthese, zodat veren uit prille leeftijd kuikens dieetgeschiedenis integreren tijdens de-chick opfokperiode als veren te vervangen natal neer [17], [18], [19]. Isotopische mengen modellen gebruiken geometrische of Bayesiaanse procedures dierlijke diëten te reconstrueren op basis van de δ 13C en δ 15N waarden weefsels consument en isotopisch afzonderlijke voedselbronnen [20], [21]. SIA en isotopische mengen modellen hebben het potentieel om relatief niet-invasieve en kosteneffectieve kwantitatieve ramingen van seabird diëten groot deel van hun jaarlijkse cyclus [22], [23], [24]. Er zijn beperkingen aan het gebruik van SIA te zeevogel diëten te kwantificeren. Wanneer de isotopische handtekeningen van prooien die eenzelfde trofisch niveau overlap bezetten, zoals in foeragevissen, overlappen, kan het moeilijk zijn om de relatieve bijdrage schatten consument diëten [25], [26]. Isotopische mengen modellen zijn alleen nuttig als de gegevens die ingaan, die een voorafgaande kennis van mogelijke prooien bronnen en hun karakteristieke waarden isotopen [15]. In veel gevallen is voorafgaande informatie ontbreekt en alle mogelijke prooi bronnen kan niet gemakkelijk worden geïdentificeerd [24]. Wanneer alle prooi isotoop waarden niet beschikbaar zijn, "vertegenwoordiger" soorten worden vaak gebruikt of meerdere bronnen worden gecombineerd a priori In deze studie we tegelijkertijd de-chick opfokvoer samenstelling van sympatrisch fokken zeevogels te kwantificeren, de Chinstrap ( Pygoscelis antarctica Onze primaire doelstellingen zijn:. 1) Gebruik gelijktijdige ophaling van SCA en SIA te vergelijken het vermogen van deze twee methoden om interjaarlijkse en inter-specifieke verschillen in dieet samenstelling Pygoscelis Materialen en methoden Ethics statement Animal gebruik in deze studie werd uitgevoerd onder goedgekeurde gebruik van dieren protocollen van de Universiteit van Californië in San Diego Institutional Animal Care en gebruik Comite (S05480) en in overeenstemming met Antarctic Conservation Act vergunningen verstrekt door de Amerikaanse National Science Foundation om S. Emslie (2006-001) en R. Holt (2008-008). maaginhoud, veren en prooi monsters Het veldwerk vond plaats in januari en februari van 2008 en 2009 een kolonie van ongeveer 4500 broedparen van Chinstrap pinguïns en 800 broedparen van Gentoo pinguïns in Cape Shirreff. We verzamelden maaginhoud monsters tijdens de-chick opfokperiode na kuikens de crèche podium (> 2,5 weken oud) had bereikt. We proefden 2-5 unieke fokkerij volwassenen terugkomst van foerageren reizen tussen 15: 00-17: 00 uur plaatselijke tijd op 5-7-dagen intervallen voor een totaal van 10-14 Gentoo pinguïns en 30 Kinbandpinguïns elk jaar. We gebruikten de water-offloading techniek naar aanleiding van een wijziging van de CCAMLR Ecosystem Monitoring Program (CEMP) Standard Methods [37]. In het bijzonder, hebben we niet de volledige inhoud van de maag te analyseren; verliepen we ongeveer de helft (ongeveer 350 g). Het grootste deel van het voedsel onder dit bovenste deel is zwaar verteerd en is het moeilijk om objectief te scheiden van prooidieren en de opname kan vooringenomenheid zowel prooi identificatie en dieet samenstelling schattingen [10], [38]. Verder hebben we deze sampling methode te rechtvaardigen als ouders gewoonlijk niet hun hele voedselketen lading te voeren aan de kuikens [39], [40]. Overmaat vloeistof werd uit elk monster maag verwijderd door zeven het door een fijne zeef weeg een monstermassa (nat gewicht) te verkrijgen. Uit deze monsters, bepaalden we het percentage van krill, vis en ander materiaal frequentie voorkomen en gewicht. We herstelde visotolieten uit de voeding monsters door wervelende monsters in een donkere bodem pan en geïdentificeerd otolieten tot het laagst mogelijke taxonomische niveau met behulp van een interne referentie collectie en een gepubliceerde otoliet gids [41]. We berekenden de frequentie voorkomen en het minimum aantal personen (MNI) van elke vis taxa volgens standaard methoden [42]. Speciaal, we geschat MNI door het optellen van het hogere aantal van links of rechts otolieten met de helft van het aantal geërodeerde Otoliths van onbekende kant om een conservatieve schatting van de totale MNI vertegenwoordigd in elk maag monster [42] bieden. Daarnaast gebruikten we otoliet metingen en gepubliceerd regressievergelijkingen in totaal en percentage totale opgeloste massa voor elke vis taxa geïdentificeerd (tabel S1) uitrekenen [7], [13], [41], [43]. Vanwege het hoge aantal kleine Pleuragramma antarcticum In februari van elk jaar, verzamelden we drie borst veren van een aselecte steekproef van 18-20 jonge kuikens van elke soort, terwijl ze waren bezig om hun geboorte koloniën vertrekken naar de zee bij 7-10 weken oud. Van 2005 tot 2009 hebben we verzamelde representatieve steekproeven van pinguïn prooidieren tijdens sleepnetten uitgevoerd langs de South Shetland Islands en noordelijk Antarctisch schiereiland en hield monsters bevroren voorafgaand aan analyse. We verder aangevuld dit prooi bibliotheek met gepubliceerde isotopen waarden van twee vissen prooi, protomyctophum bolini Kopen en Champsocephalus gunnari We schoongemaakt veren met behulp van een 02:01 chloroform: methanol spoeling, de lucht gedroogd en snijd ze in kleine fragmenten met roestvrijstalen schaar. We gehomogeniseerd geheel prooi monsters, droogde ze gedurende 48 uur in een oven bij 60 ° C en daarna geëxtraheerd lipiden uit deze monsters met behulp van een Soxhlet apparaat met 01:01 petroleumether: Ethyl-Ether oplosmiddel mengsel gedurende 8 uur [46]. We flash-verbrand (Costech ECS4010 elementaire analyser) ongeveer 0,5 mg van elk veer en prooi monster geladen in tinnen kopjes en geanalyseerd op koolstof en stikstof isotopen (δ 13C en δ 15N) via een interface Thermo Delta V Plus continue stroom stabiele isotoop ratio massaspectrometer (CFIRMS). Raw δ waarden werden genormaliseerd op een twee-puntsschaal met behulp van glutaminezuur referentiematerialen met lage en hoge waarden (dwz USGS-40 (δ 13C = -26,4 ‰, δ 15N = -4,5 ‰) en USGS- 41 (δ 13C = 37,6 ‰, δ 15N = 47,6 ‰)). Sample precisie op basis van herhaalde monster en naslagmateriaal was 0,1 ‰ en 0,2 ‰ voor δ 13C, en δ 15N, respectievelijk. stabiele isotoop ratio's zijn uitgedrukt in δ notatie in promille eenheden (‰), volgens de volgende vergelijking: Waar X 13C of 15N en R is de overeenkomstige verhouding 13C / 12C of 15N / 14N. De R We gebruikten vier modelvarianten van de SIAR Bayesiaanse mengen model [21] in de R omgeving (R Development Core Team 2007) ons vermogen om kuiken dieet samenstelling (tabel S2) te kwantificeren te verkennen. De SIAR modelschattingen kansverdelingen van meerdere bronnen bijdragen aan een mengsel, terwijl de boekhouding voor de waargenomen variabiliteit in de bron en mengsel isotopische handtekeningen, dieet isotopische fractionering, en elementaire concentratie. We gebruikten twee SIAR model varianten met twee prooi bronnen (Antarctisch krill vs. "vis") op een dieet samenstelling voor elke soort /jaar combinatie met de δ 13C en schatten δ 15N waarden van chick veren. Model 1 maakt gebruik van de δ 13C en δ 15N waarden van een representatieve vissoorten, P. antarcticum We gebruikten twee andere varianten van de SIAR mengen uitvoering met meerdere bronnen prooi (6-7 afhankelijk pinguïns soorten) op werkwijzen voor het integreren van maaginhoud gegevens naar de taxonomische betere opheldering verdere beoordeling samenstelling van de vis deel van de pinguïn diëten. Voor deze modellen beperkt we onze analyses veer gegevens kuiken van 2008, toen de vis portie chick diëten was de meest uiteenlopende. Model 3 is een eerste multi-source model schatten van de relatieve bijdrage van krill ( E. Superba Statistische analyse De statistische berekeningen werden uitgevoerd met behulp van SAS (versie 9.1). We analyseerden SCA gegevens om te testen op verschillen tussen jaren en soorten gebruik van aparte gegeneraliseerde lineaire modellen (Proc Genmod). We gebruikten een binomiale fout distributie en logit koppeling functie voor gegeneraliseerde lineaire modellen met de procentuele samenstelling (door natte massa) of de frequentie optreden van elk van onze drie belangrijkste prooi groep (krill, vis, en 'andere' prooien) als responsvariabelen. Voor modellen die MNI gebruikt in plaats van vis en bereide vis massa per monster als responsvariabelen, gebruikten we een Poisson-verdeling fout met een logit koppeling functie. Voor alle gegeneraliseerde lineaire modellen voerden we post-hoc analyses met behulp van een Bonferroni correctie en gerapporteerd chi-kwadraat en p-waarden uit de likelihood ratio testen statistieken voor type 3-tests. Om te testen voor verschillen in het kuiken veer δ 13C en δ 15N waarden gebruikten we multivariate variantie analyse (MANOVA) samen met Tukey-Kramer Multiple vergelijking waarden tussen soorten en jaren met behulp van PROC ANOVA. We gebruikten een vergelijkbare MANOVA met de δ 13C en onderzoeken δ 15N waarden van soorten in onze prooi bibliotheek. We gebruikten model 95% geloofwaardigheid intervallen schattingen van krill vs. vis te vergelijken tussen twee-source SIAR modelvarianten (Modellen 1 en 2) en SCA natte massa, en het percentage aandeel van de afzonderlijke vissoorten vissen deel van chick diëten onder de multi- bron SIAR modelvarianten (Modellen 3 en 4) en SCA otoliet-afgeleide opgelost vis massa. Om directe vergelijking tussen SIAR modellen en SCA te vergemakkelijken, berekenden we Bayesiaanse gemiddelden en 95% geloofwaardigheid intervallen voor iedere SCA dataset met behulp van Markov keten Monte Carlo (MCMC) simulaties via Winbugs (versie 1.4). Deze MCMC simulaties werden uitgevoerd met behulp van de niet-informatieve Dirichlet voorafgaand door een identiek aantal iteraties, dunner, en teruggooi als onze SIAR model analyse. Daarnaast gebruikten we Chi-kwadraat goedheid van fit tests om de verdeling van de gemiddelde geschatte percentage bijdrage van de afzonderlijke prooivissoorten om voeder bij meerdere bronnen SIAR modellen en varianten SCA gegevens te vergelijken. De gegevens werden onderzocht voor normaliteit en gelijke variantie, alle tests waren tweezijdig en betekenis werd aangenomen op het niveau van 0,05. Stabiele isotoop waarden van kuiken veren en prooidieren worden gepresenteerd ± standaarddeviatie (SD), terwijl de voeding samenstelling schattingen van maaginhoud analyse worden gepresenteerd ± standaardfout (SE) in tabellen en ± 95% geloofwaardigheid intervallen in cijfers. maag inhoudsanalyse Chinstrap pinguin maag monsters had een hoger percentage bijdrage van krill ten opzichte van Gentoo penguin monsters (tabel 1a; χ 2 1 = 10.91, p = 0,0010). Echter, we vonden geen verschillen op jaar (χ 2 1 = 0,22, p = 0,6375) of een soort * jaar interactie (χ 2 1 = 0,00, p = 0,9805). Op dezelfde manier, Gentoo penguin monsters bevatten een aanzienlijk hoger percentage bijdrage van de vis ten opzichte van Chinstrap pinguïn monsters, (χ 2 1 = 12,24, p = 0,0005), maar we konden geen verschillen tussen de jaren (detecteren χ 2 1 = 0,08, p = 0,7755) of een soort * jaar interactie (χ 2 1 = 0,26, p = 0,6078). De procentuele bijdrage aan de maag monsters van andere prooidieren, met inbegrip van koppotigen, vlokreeftjes en andere euphausiid soorten verschilde niet door pinguïnspecies (χ 2 1 = 0,00, p = 0,9694), jaar (χ 2 1 = 0,36, p = 0,5468) of een soort * jaar interactie (χ 2 1 = 0,00, p = 0,9694). We vonden bewijs van krill in alle Chinstrap penguin samples en in alle, maar een Gentoo penguin monster (Tabel 1a). We vonden bewijs van vissen in alle Gentoo penguin samples en 36,7-50,0% van Chinstrap pinguïn monsters, zelfs wanneer er geen waarneembare natte massa van vis (Tabel 1a). Echter, de frequentie van voorkomen van vis in Chinstrap pinguin monsters niet verschillen tussen jaren (χ 2 1 = 1,09, p = 0,2966). Ook heeft de frequentie voorkomen van andere prooidieren niet verschillen van Penguin soort (χ 2 1 = 1,57, p = 0,2107), jaar (χ 2 1 = 0,00, p = 0,9481 ), of een soort * jaar interactie (χ 2 1 = 0,00, p = 0,9481). De MNI van vis en bereide vis massa per monster verschilde tussen soorten en jaren (tabel 1b ). Stormbandpinguïn diëten hadden lagere MNI en bereide vis massa dan Gentoo pinguïns (MNI: χ 2 1 = 959,14, p < 0,0001; opgeloste massa: χ 2 1 = 959,14, p < 0,0001 ). Across soorten en jaren Kinbandpinguïns hadden hogere opgeloste vis massa in het dieet van de monsters in 2008 ten opzichte van 2009, terwijl de Gentoo pinguïns beide onderste MNI en bereide vis massa's hadden in 2006 ten opzichte van 2009 (MNI: χ 2 1 = 147,74 , p < 0,0001; opgeloste massa: χ 2 1 = 1122,46, p < 0,0001). Een totaal van 96,3% van alle otolieten waren herkenbaar aan ten minste het geslacht niveau, met zes en vijf vissen taxa vertegenwoordigd in Chinstrap en Gentoo pinguïn diëten monsters, respectievelijk (Tabel S1). We hebben δ 15N en δ 13C waarden van pinguïnkuiken veren verschilde per soort (Wilks 'λ, p < 0,0001), jaar (Wilks' λ, p = 0,0409) en had een belangrijke soort * jaar interactie (Wilks 'λ, p < 0,0001). Gentoopinguïn kuikens hadden hogere veren δ 15N waarden dan Chinstrap pinguïn kuikens in beide jaren (tabel 2, afb. 1). Echter, terwijl Gentoo penguin chick veer δ 15N-waarden waren hoger in 2009 ten opzichte van 2008, Chinstrap penguin chick veer δ 15N waarden verschilde niet tussen jaren. Chinstrap en Gentoo pinguïnkuikens had soortgelijke veren δ 13C waarden in 2008, maar lagere en hogere waarden voor Chinstrap en Gentoo pinguïns in 2009, respectievelijk (tabel 2). We vonden δ 15N en δ 13C waarden van soorten in onze bibliotheek van gemeenschappelijke pinguïn prooien ook aanzienlijk verschilde (Wilks 'λ, p < 0,0001). De δ 15N en δ 13C waarden verschilden sterk tussen krill en vissoorten, terwijl isotoop waarden overlapt onder vele vissoorten (tabel 2, afb. 1). De twee-source SIAR model variant die gebruikt P. antarcticum Multi-source SIAR modellen Beide multi-source SIAR modelvarianten (modellen 3 en 4) voorspelde dat Antarctisch krill bestond uit de grootste prooi component van Chinstrap en Gentoo penguin chick diëten in 2008 (tabel 4). Daarnaast zowel multi-source SIAR modellen in grote lijnen met twee-source SIAR model schattingen van het relatieve aandeel van krill vs. overeengekomen alle vissoorten opgeteld (tabellen 3 en 4). Echter, onze eerste multi-source SIAR model (Model 3) had moeite met het inschatten van het relatieve aandeel van de afzonderlijke vissoorten zowel pinguïnspecies chick diëten in 2008. SIAR Model 3's 95% geloofwaardigheid intervallen breed overlapt over vissoorten en de gemiddelde relatieve aandeel van de elke vissoort verschilde van schattingen met behulp van otoliet opgeloste massa (Tabel 4, figuur 3; Chinstrap. χ 2 5 = 62,65, p < 0,0001; Gentoo: χ 2 4 = 41,70, p <. 0,0001) In tegenstelling, de a posteriori Discussie maag inhoudsanalyse Onze SCA analyse wijst op een aantal van de mogelijke vertekeningen die inherent zijn bij het gebruik van deze methode. Net als bij eerdere studies bij Kaap Shirreff, zagen we het bewijs van vissen zoals otolieten, een weegschaal en lenzen in veel Chinstrap penguin monsters, zelfs wanneer er geen meetbare hoeveelheid vis weefsel door natte massa [34], [36]. Dit bewijs suggereert dat vis biomassa verbruikt door volwassenen verteert volledig voorafgaand aan hun terugkeer naar de broedkolonie of, waarschijnlijker, wordt geleverd aan kuikens in de zwaar-verteerde component van de volwassen maaginhoud die niet objectief kan worden gekwantificeerd [6], [10] . Verder omdat we maag monsters in de late namiddag verzameld, onze monster is exclusief volwassenen die 's nachts foraged en doorgaans een veel hoger percentage en voorkomen van vis in de maag monsters [34], [48] hebben. Bovendien voersamenstelling schattingen afgeleid van SCA in onze studie waren vaak zeer variabel, waardoor het moeilijk om verschillen tussen jaar pinguïnspecies (tabel 1) te detecteren. Deze bevinding lijkt niet uniek in zeevogel voedingskundige studies te zijn met behulp van SCA, die vaak vereist hoge steekproefomvang en de grote verschillen tussen de groepen om inter-jaarlijkse of species-specifieke verschillen in dieet samenstelling te detecteren [3], [9]. Echter, onze studie suggereert dat de analyse van otolieten nog steeds gedetailleerde informatie over de soortspecifieke en temporele variatie in de consumptie van vis prooidieren als totale voeding samenstelling schattingen afgeleid van maaginhoud natte massa zijn minder informatief. twee-source SIAR modellen voorspelden een relatief grotere bijdrage van vis tot kuiken opvoeding diëten in beide soorten in vergelijking met de ramingen van SCA biomassa. Dit resultaat is niet onverwacht, aangezien SCA wordt beschouwd als de hoeveelheid vis in de voeding van deze soorten onderschat door de vertering en diel biases bovenstaand [10] beschreven, [12], [48]. Bovendien, twee-source SIAR modellen ontvangen ook het minst variabele voorspellingen van voedersamenstelling in vergelijking met SCA. De SIA kuiken veren voorzien een gemiddelde waarde van het dieet van elk individu kuiken gedurende de tijd van de veer groei in de chick-opfokperiode [18], [19]. Daarentegen SCA gegevens vormen een reeks van "snap-shots" (in dit onderzoek om de 5 tot 7 dagen) van het voedsel dat een van beide ouders voeden haar chick [3]. Onze studie suggereert dat SIA van weefsels die diëten gedurende lange perioden te integreren zijn aangeboren minder variabel dan SCA gegeven een vergelijkbare steekproefgrootte en zijn meer geschikt voor de behandeling van jaar tot jaar verschillen in chick diëten. Bijvoorbeeld, de twee-source SIAR modellen die in onze studie konden identificeren interjaarlijkse en soort-specifieke verschillen in de relatieve abundantie van vis en krill in diëten niet meteen duidelijk behulp SCA. Wanneer vooraankondiging op prooi soortensamenstelling is beperkt, zoals buiten het broedseizoen, met behulp van een vertegenwoordiger prooi bron in isotoop mengen modellen kunnen belangrijke informatie verschaffen over zeevogel diëten wanneer weinig anders bekend [24]. Echter, onze resultaten suggereren ook dat variatie in prooi soortensamenstelling binnen trofische of functionele groepen kunnen significante verschillen in dieet samenstelling die niet uit isotoop waarden of het mengen van modelvoorspellingen met behulp van enige vertegenwoordiger prooi bronnen zou maskeren. Dit resultaat was het meest duidelijk bij het onderzoek van het effect van vissen prooi δ 15N waardes op kuiken veer δ 15N waarden en de twee-source isotoop mengen modellen die gebruikt worden in onze studie. Bijvoorbeeld, wist Chinstrap penguin chick veer δ 15N waarden niet verschillen tussen de jaren (tabel 2). Bovendien, de 95% interval geloofwaardigheid van dieet schatting van de twee-source SIAR model met P. antarcticum bij de parametrering van onze twee multi-source SIAR modellen die we gebruikt otoliet gegevens naar de juiste vissen prooi bronnen te selecteren in elke soort model (tabellen S1 en S2) op te nemen. Echter, onze eerste multi-source SIAR model (Model 3) had moeite juist inschatten van de individuele soorten samenstelling van de vis deel van de pinguïn diëten als gevolg van de algemene overeenkomsten in δ 13C en δ 15N waarden bij veel van de vissoorten opgenomen als prooi bronnen (afb. 3). Antarctische vissoorten in het algemeen consumeren krill en andere vissoorten en als gevolg van hun soortgelijke tropische niveau, deze vissoorten hebben de neiging om soortgelijke δ 15N waarden [45] hebben.
) en Gentoo ( P. Papua
) pinguïns en hoogtepunt methoden voor het integreren van SCA gegevens om de juistheid van verhogen dieet samenstelling schattingen met behulp van SIA. ramingen van SCA biomassa waren zeer variabel en onderschat het belang van soft-bodied prooien zoals vis. Twee-source, isotoop mengen modelvoorspellingen waren minder variabel en geïdentificeerd inter-jaar- en soortspecifieke verschillen in de relatieve hoeveelheden vis en krill in pinguïn diëten niet meteen duidelijk met behulp van SCA. In contrast, multi-source isotoop mengen modellen hadden moeite met het inschatten van de voedingskundige bijdrage van vissoorten bezetten vergelijkbare trofische levels zonder verfijning met behulp van SCA afgeleid otoliet data. Over het algemeen, ons vermogen om te volgen interjaarlijkse en soortspecifieke verschillen in penguin diëten met behulp van SIA werd versterkt door de integratie van SCA data aan het mengen modes isotoop op drie manieren: 1) het selecteren van geschikte prooi bronnen, 2) weging combinaties van isotopen soortgelijke prooi in twee -source mengen modellen en 3) raffinage voorspelde bijdragen van de isotoop soortgelijke prooi in multi-source-modellen
voor elk trofisch of functionele groep [24], [27], [28]. Bovendien, terwijl de studies van de zeevogel diëten gebruik van SIA worden steeds gemeengoed, weinig studies vergeleken gelijktijdige kwantitatieve ramingen van het dieet samenstelling tussen SCA en SIA [28], [29]. Bovendien is het ook gebruikelijk om SIA-gegevens te vergelijken SCA prooi frequentie van voorkomen in plaats van geschiktere massa gebaseerde schattingen van voedersamenstelling afgeleid van SCA [30], [31], [32].
) en Gentoo penguin ( P. papua
) over twee fokken seizoenen bij Kaap Shirreff, Livingston Island, Antarctica (62 ° 28 ', 60 ° 46'W) met behulp van zowel SIA en SCA. Net als bij andere Antarctische zeevogels, Pygoscelis
pinguin diëten zijn over het algemeen samengesteld uit zoöplankton, hoofdzakelijk Antarctisch krill ( Euphausia superba
), en soft-bodied, hogere trofische prooidieren, zoals vis [ ,,,0],33]. As-chick grootbrengen diëten zijn goed bestudeerd met behulp van SCA op deze site, biedt een uitstekende case study voor de vergelijking met SIA [34], [35], [36]. Wij streven naar een beter begrip van de relatieve voordelen van beide methoden en benadrukken het gebruik van SCA aan isotopen mengen modellen informeren beter te kwantificeren van de voeding van zeevogels gebruik SIA
pinguïn kuikens, 2) vergelijken de voorspellend vermogen van een twee-bron op te sporen (krill vs. vis) lineair mengen model onder degenen met behulp van een representatieve vissoorten en die met behulp van een a priori
gemiddeld soort en jaar-specifieke vis waarden, en 3) het evalueren van een methode van a posteriori
integratie van SCA gegevens naar de taxonomische samenstelling van de vis gedeelte van diëten met behulp van een multi-source Bayesiaanse mengen model beter toe te lichten.
otolieten, maten we een willekeurige sub-steekproef van 20-75 P. antarcticum
otolieten per monster en gebruikte deze waarden gereconstitueerde massa te schatten voor deze soort.
[44], [45].
stabiele isotoop analyse
standaard waarden waren gebaseerd op de Weense PeeDee Belemnite (VPDB) voor δ 13C en atmosferische N 2 voor δ 15N.
Isotopen mengen modellen
, die gewoonlijk wordt gevonden in de Pygoscelis
pinguin diëten als de "vis" bron [6]. Model 2 maakt gebruik van soorten en jaar specifieke "vis" δ 13C en δ 15N waarden berekend door het gemiddelde van de δ 13C en δ 15N waarden van meerdere vissoorten gewogen op basis van hun relatieve procent bereide vis massa (tabellen S1 en S2).
) en alle vissoorten in onze prooi bibliotheek geïdentificeerd uit otolieten in elke soort 'maaginhoud (tabellen S1 en S2). Model 4 is een a posteriori
geïnformeerde model waarbij we beperkt het resulterende posterior vestigt op die waarin het relatieve belang van individuele vissoorten werd gerangschikt in overeenstemming met de overvloed van elk geïdentificeerd door middel van otoliet analyse soorten. Voor Model 4, beperkt we posterior trekt om alleen die waarbij de geschatte proportionele bijdrage van de meest voorkomende vissen prooi op basis van bereide vis massa van meer dan de geschatte proportionele bijdrage van de tweede meest overvloedige vis prooi was, en voor de tweede meest voorkomende vissen prooi groter dan de derde meest voorkomende en zo verder voor alle vissoorten. Voor zowel de eerste (Model 3) en hoogte (Model 4) multi-source-modellen, we vatte ook resultaten over vissen prooi en schatte de proportionele bijdrage van elke vissoort om de vis deel (dat wil zeggen met uitzondering van krill) van de pinguïn diëten. Voor alle SIAR modellen opgenomen we Pygoscelis
penguin veer δ 15N en δ 13C discriminatie factoren [47] en liep 1.000.000 iteraties, verdund met 15, met een initiële zich ontdoen van de eerste 40.000 resulterende in 64.000 posterior trekt.
Resultaten
isotopische handtekeningen van chick veren en prooi
Twee-source SIAR modellen
isotoop waarden als een vertegenwoordiger "vis" bron (model 1) en de variant die een jaar en soortspecifieke gewogen "vis" isotopen die worden gebruikt (Model 2) beide voorspelde dat Gentoo pinguïnkuikens verbruikt relatief minder krill en nog veel meer vis dan Kinbandpinguïns in beide jaren (tabel 3). Echter, bij de behandeling van model 95% geloofwaardigheid tussenpozen deze twee model varianten verschilden in hun vermogen om soortspecifieke, inter-jaarlijkse verschillen in dieet samenstelling te detecteren. Hoewel beide twee-source SIAR modelvarianten voorspelde dat Gentoo penguin chick diëten bevatten een hoger percentage van krill in 2008, slechts Model 2 ontdekte een grotere hoeveelheid vis in Chinstrap penguin chick diëten in 2008 ten opzichte van 2009 (tabel 3). Twee-source SIAR modelvarianten voorspelde een hogere bijdrage van de vis in het kuiken diëten van beide pinguïnspecies in vergelijking met voeding samenstelling schattingen afgeleid van SCA natte massa (fig. 2). SCA schattingen werden ook meer variabel dan SIAR modelvoorspellingen voor Gentoo penguin chick diëten. Bovendien SCA afgeleide schattingen van de gemiddelde bijdrage van krill en vis in diëten beide species vielen buiten onze twee-source SIAR modellen 95% bovenste en lage geloofwaardigheid tijdstippen (Fig 2;. Tabellen 1 en 3).
geïnformeerde meerdere bronnen SIAR model (model 4) beter dan de eerste multi-source SIAR model (model 3) een raming van de soortensamenstelling van de vis gedeelte van chick diëten. Terwijl Model 4's voorspelling van de gemiddelde relatieve aandeel van elke vissoort in Chinstrap penguin chick diëten enigszins afwijken van schattingen van otoliet opgeloste massa (χ 2 5 = 14.55, p = 0,0125), de resulterende 95% geloofwaardigheid intervallen daalden met 53,5 ± 17,2% ten opzichte van Model 3 (uitersten: 33,2-82,2%; tabel 4, afb. 3). Bovendien Model 4 voorspelling's van de gemiddelde relatieve aandeel van elke vissoort in diëten Gentoo pinguïnkuikens 'was gelijk aan de ramingen van otoliet opgeloste massa (χ 2 4 = 3,40, p = 0,4949). Daarnaast werden de resulterende 95% geloofwaardigheid intervallen verminderd met 52,0 ± 27,7% ten opzichte van Model 3. (Bereik:. 17,4-76,2%; Tabel 4, Figuur 3)
twee-source, SIAR modellen
als de vis prooi bron (Model 1) tussen de jaren overlappen (tabel 3). In tegenstelling tot 95% geloofwaardigheid tussenpozen van twee-source SIAR model met behulp van jaar- en soortspecifieke gewogen "vis" waarden (Model 2) suggereren een grotere overvloed aan vis in Chinstrap penguin chick diëten in 2008 ten opzichte van 2009, dat werd bevestigd door otoliet afgeleid, gemiddelde opgeloste vis massa. In 2008, de vis gedeelte van Chinstrap penguin chick diëten bestond uit zes vissoorten met een geschatte δ 15N waarde van 7,9 ± 0,7 ‰, terwijl de P. antarcticum
(δ 15N: 9,4 ± 0,5 ‰) was de enige vissoorten in 2009 diëten (tabellen S1 en S2). Hoewel dit 1,5 ‰ verschil is klein in verhouding tot 4,6-6,1 ‰ verschillen tussen vis en krill, het was genoeg om inter-jaarlijkse vergelijkingen van Chinstrap penguin chick diëten beschamen in onze studie.
Multiple-source, SIAR modellen
 De meest voorkomende diarree-trigger (die je negeert)
Een paar jaar geleden had ik een cliënt met colitis ulcerosa die eindelijk het idee bevestigde dat onze levensstijl, en meer specifiek stress, het enige kan zijn dat ons ervan weerhoudt om gezond te w
De meest voorkomende diarree-trigger (die je negeert)
Een paar jaar geleden had ik een cliënt met colitis ulcerosa die eindelijk het idee bevestigde dat onze levensstijl, en meer specifiek stress, het enige kan zijn dat ons ervan weerhoudt om gezond te w
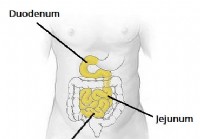
 SIBO (Small Intestinal Bacterial Overgrowth) Oorzaken, symptomen, behandelingen en dieet
Definitie en feiten van bacteriële overgroei in de kleine darm (SIBO) Afbeelding van SIBO (Small Intestinal Bacterial Overgrowth) SIBO staat voor bacteriële overgroei in de dunne darm. SIBO kan wor
SIBO (Small Intestinal Bacterial Overgrowth) Oorzaken, symptomen, behandelingen en dieet
Definitie en feiten van bacteriële overgroei in de kleine darm (SIBO) Afbeelding van SIBO (Small Intestinal Bacterial Overgrowth) SIBO staat voor bacteriële overgroei in de dunne darm. SIBO kan wor
 Waarom krijg je galstenen?
Galstenen zijn gemaakt van materiaal in de gal dat kristalliseert. De exacte oorzaak van galstenen kan meestal niet worden vastgesteld. Enkele mogelijke oorzaken zijn: Hoog cholesterol in de gal:
Waarom krijg je galstenen?
Galstenen zijn gemaakt van materiaal in de gal dat kristalliseert. De exacte oorzaak van galstenen kan meestal niet worden vastgesteld. Enkele mogelijke oorzaken zijn: Hoog cholesterol in de gal: