Kjennetegn på Epstein Barr-virus varianter assosiert med mage carcinoma in Southern Tunisia
Abstract
bakgrunn
EBV-assosiert magekarsinom (EBVaGC) har en distinkt kliniske funksjoner og dens utbredelse er variabel over hele verden.
Resultater
for å fastslå utbredelsen av EBVaGC i Tunisia, ble EBV-kodet små RNA (EBER) uttrykk vurdert i 81 gastrisk karsinom (GC) prøver. Atom EBER uttrykket ble påvist i 12 av 81 GC tilfeller (14,81%) og samstemmighet mellom poengsum utvalg Eber flekker og antall EBV DNA-kopier som estimat av QPCR er observert. På den annen side har vi funnet at EBVaGC sterkt korrelert med alder ved diagnose, og svakt med tumor differensiering og venøs invasjon.
Videre ble EBVaGC prøvene utsatt for å bestemme de EBV DNA-polymorfismer. Våre resultater viser en unik genetisk profil av EBV-stammene om A- og D-type, F prototype, retensjonen av Xho
-restriksjonssete, og den 30 bp del-lmp1 variant. Ifølge våre tidligere studier på nasopharyngeal karsinom (NPC), foreslo vi at EBV stammer knyttet til GC og NPC delte noen likheter i tunisiske pasienter.
Konklusjon
Forekomsten av EBVaGC er av 14,81% i den sørlige Tunisia og at felles EBV belastningen er assosiert med både NPC og GC som sannsynligvis vil avvike fra asiatiske stammer. Våre funn støtter derfor en viss geografisk spredning av EBV stammer som ikke er begrenset til EBV-assosiert malignitet.
Nøkkelord
magekarsinom EBV EBER polymorfismer Bakgrunn
Gastric karsinom (GC) er den nest største årsaken til kreftdød verdensomspennende [1]. Forekomsten av GC varierer fra et geografisk område til et annet, noe som tyder på at genetiske og miljømessige faktorer, inkludert Helicobacter pylori-infeksjon
anses for å bidra til gastrisk carcinogenesis [2, 3]. I Sør-Tunisia, den årlige forekomsten av GC varierer fra 2,6 til 4,8 /100 000 personer [4]. I de få siste årene, har mange rapporter utforsket sammenhengen mellom Epstein-Barr-virus (EBV) infeksjon og GC [5-9]. Den EBV-assosierte GC (EBVaGC) har blitt vist ved nærværet av ensartet ekspresjon av EBV-kodet liten RNA (EBER) i GC-celler [10], påvisning av monoklonale EBV episomer i GC-celler [11] og økningen av serum antistoffer mot virale kapsid-antigen [12]. Forekomsten av EBVaGC varierer 2-18% som er rapportert av tidligere studier [13-20]. Videre kliniske kjennetegn EBVaGC inkluderer mannlig dominans, relativt yngre alder og plassering i den proksimale magen [9, 21]. Den EBVaGC viser en lavere rate av lymph node-engasjement og har en forholdsvis gunstig prognose i forhold til EBV-negative on [22].
I EBVaGC, er infeksjon av tumorceller, karakterisert ved en type I-mønster av ventetid, i hvilken uttrykk av virale latente gener er begrenset til Epstein-Barr kjernekraft antigen (EBNA) -1, Eber latent membranprotein LMP-2A og transkripsjoner fra Bam
HI A mot høyre ramme (BARF) -0 og -1 [11, 23]. Siden EBV produkter av type I latent infeksjon har blitt vist å være involvert i EBV-mediert tumorogenesis, ble det foreslått at det onkogene prosessen i EBVaGC kunne drives ved forskjellige mekanismer fra de av konvensjonelle GC. Deretter kan EBVaGC malignitet betraktes som en annen enhet blant GC som krever større oppmerksomhet for å bedre pasientbehandlingen.
Studien ble gjennomført med sikte på å fastslå utbredelsen av EBVaGC i sørlige Tunisia og senere for å analysere spesifikke EBV polymorfismer i svulsten isolerer.
Materiale og metode
Pasientens egenskaper, En totalt 81 primær mage karsinom ble samlet inn, mellom januar 1999 og desember 2009 fra pasienter som gjennomgikk radikal kirurgisk reseksjon ved Institutt for Digestive Surgery av Habib Bourguiba universitetssykehus (Sfax, Tunisia). Alle pasienter ga informert samtykke før prøvetaking i henhold til institusjonelle retningslinjer. Ingen av pasientene hadde pre-operative eller postoperativ kjemoterapi. Klinisk-patologisk parametere som kjønn, alder, anatomiske området, histologisk type patologisk stadium, tumorstørrelse og venøs invasjon ble evaluert ved å gjennomgå medisinske diagrammer og patologiske poster. På tidspunktet for kirurgi, i en alder av pasientene varierte fra 18 til 94 år (gjennomsnitt: 59,58 år). Den anatomiske stedet for tumoren ble bestemt i henhold til den dominerende lokalisasjon som cardia (n = 8), kropp (n = 22), og antrum (n = 48). De histologiske subtyper ble klassifisert i henhold til kriteriene for Lauren [24] som intestinal type (n = 47) og diffus type (n = 34), men også av Verdens helseorganisasjon som dårlig differensiert (n = 43), moderat differensiert (n = 26) og godt differensiert (n = 9). Den kliniske fasen av sykdommen ble bestemt i henhold til svulsten, node og metastase (TNM) klassifisering av det amerikanske Joint Committee on Cancer [25]. Vår kohort inneholder 9 pasienter i stadium I eller II og 47 pasienter med stadium III eller IV.
In-situ hybridisering
EBV ble identifisert av uttrykket av EBV-kodet små RNA (EBER). Kort, in situ
hybridisering (ISH) Analysen ble utført på 3 mikrometer parafin-blokkseksjoner med EBV oligonukleotidprober komplementære til EBER-1 og -2 i henhold til produsentens instruksjoner (PNA ISH deteksjon kit, Dako Cytomation). De hybridisering signaler ble visualisert med diaminobenzidine (DAB) og positiv atom signal ble anerkjent som mørk brun atom flekker under lett mikroskopi. Som positiv kontroll, ble del fra en kjent EBER-positive NPC vev som brukes. Farging ble scoret 1-3 etter andelen av farget cellekjerner i vevssnitt. Score 3 ble tilskrevet da positivt signal ble observert i mer enn 75%, ballen 2 på 75 til 50% og scorer 1 på mindre enn 50% av cellene.
DNA-ekstraksjon
DNA ble ekstrahert fra formalinfiksert, parafin innebygde vev. Etter svulst identifikasjon på Hematoxylin-eosin-farget lysbilder, ble tumor områder skrapt fra 40 mikrometer tykke parafinsnitt. De oppsamlede materialer ble av-vokset ved vasking i xylen og skyllet i etanol. Tørkede vev ble fordøyd med proteinase K i nærvær av SDS ved 55 ° C over natten, etterfulgt av fenol /kloroform-ekstraksjon som tidligere beskrevet [26]. Mengden av DNA ble undersøkt ved spektrofotometer og lagret ved -20 ° C for videre bruk.
PCR-RFLP og sekvense
fem regioner av EBV-genomet var målrettet for polymorfisme analyse av PCR, RFLP eller sekvensering. Polymorfismene studert omfatte A eller type B i EBNA-3C-genet, C eller D-type i BamHI-W1 /I1 region, prototypen F eller f variant i BamHI-F-regionen, tap av et Xho
jeg området i første ekson av BNLF1gene (Xho
i-tap variant) og 30 bp sletting i tredje ekson av det samme genet (30 bp del-lmp1 variant). Genotyping ble utført på DNA fra GC pasienter EBV-positive og som kontroll, analyserer vi 10 DNA-prøver fra nasopharyngeal mucosa positive for EBV.
PCR-amplifikasjon ble utført på 200 ng DNA i et endelig reaksjonsblanding på 50 pl inneholdende 0,2 pM av hver primer, 200 uM dNTP, 2 mM MgCl2, 1 x PCR buffer og 1 enhet Taq DNA-polymerase (Fermentas). Primersekvensene og størrelsen av PCR-produkter er vist i tabell 1.Table en Oppsummering av primersekvensene omfatter de EBV-typer og varianter
EBV-gener og regioner
(typer eller varianter )
Primer sekvenser (5'-3 ')
Produkt størrelse (bp)
EBNA-3C genet plakater (type A eller B)
F : AGAAGGGGAGCGTGTGTTGT
R: GGCTCGTTTTTGACGTCGGC
PCR produktene
153: type A
246: type B
BamH1-W omegn (type C eller D)
F: ACCTGCTACTCTTCGGAAAC
R: TCTGTCACAACCTCACTGTC
PCR + Bam
HI fordøyelsen
205: type C
130 + 75: type D
BamH1-F omegn (f variant)
F: TCCCACCTGTTACCACATTC
R: GGCAATGGGACGTCTTGTAA
PCR + Bam
HI fordøyelsen
198: F variant
127 + 71: f variant
Exon 1 av BNLF1 genet plakater (XhoI variant)
F: ACAATGCCTGTCCGTGCA
R: AGAAACACGCGTTACTCT
PCR + Xho
jeg fordøyelsen
497: Xho
I-
340 + 157: Xho
i +
Exon 3 av BNLF1 gen product: (30bpdel-LMP1)
F: TGGAGGGAGAGTCAGTCAGGC
R: ATTGACGGAAGAGGTTGAAAAC
PCR produktene
224: 30bpdel-lmp1
254: wt-lmp1
F: Spiss primer
R: Revers primer
C /D og F /f typing er basert på BamHI
fordøyelse av hvert PCR-produkt. De enzymatiske reaksjoner ble utført i et endelig volum på 20 ul inneholdende 10 ul av PCR-produkt, 1X fordøyelse buffer og 10 enheter av BamHI
enzym (Fermentas). Etter inkubering over natten ved 37 ° C, ble produktene analysert på 2% agarosegel og visualisert ved etidiumbromidfarging, under UV-belysning. De samme forholdene som er beskrevet ovenfor, ble vurdert til å identifisere XhoI
området i første ekson av BNLF1gene. DNA fra B95.8 (GenBank tiltredelse No.V01555) og C666-1 cellelinje (GenBank aksesjonsnr ABV54173) ble anvendt som kontroll for EBV-typer og varianter. Den C666-1 cellelinje avledet fra et kinesisk NPC som havner A /C typer, f variant, Xho
I-tap variant og 30 bp del-lmp1 variant.
DNA-sekvensering ble utført på de rensede PCR produktene den tredje ekson av BNLF1 genet ved hjelp av SV gel Purification kit (Promega). Cycle sekvensering ble utført ved hjelp av ABI PRISM Big Dye Terminator Cycle Sequencing Kit (Applied Biosystems) i henhold til produsentens instruksjoner. Alle sekvenser ble utført begge veier. De sekvense Resultatene ble deretter sammenlignet med EBV sekvenser av B95.8 (GenBank tiltredelse No.V01555), kinesisk NPCer (CaO celler [27], C666-1 celler (GenBank tilgangsnummer ABV54173) og NPC10 biopsi [28] og tunisiske NPC prøver (CV4, CV5, og CV6 [29]).
sanntid kvantitativ PCR
Q-PCR-analyse rettet mot BamHI-W regionen av EBV-genom ble utført på 12 EBV-positive prøvene viser nukleær farging og 6 EBV-negative prøver. Reaksjoner ble utført på en iCycler iQ ™ real-time PCR-system (Bio-Rad), ved anvendelse av primere som flankerer BamHI-W-regionen av EBV-genomet og TaqMan probe som tidligere beskrevet [30]. Alikvoter på 1 ug DNA ble anvendt for amplifikasjon i et totalt reaksjonsvolum på 25 ul inneholdende 300 nM av hver primer, 25 nM av TaqMan probe, 1X PCR-buffer, 2 mM MgCl2, 200 pM av hver dNTP, og 1 enhet av GoTaq Hot start-polymerase (Promega). standard~~POS=TRUNC kurven~~POS=HEADCOMP ble fremstilt ved hjelp av serielle 10-gangers fortynninger av det rekombinante plasmid pGEMT /BamHI-W. Prøvene ble kjørt i duplikat, og resultatene i gjennomsnitt til å beregne EBV virusmengde uttrykt i kopier per reaksjon.
Statistisk analyse
Statistisk analyse ble utført med SPSS 13.0 statistisk program for Windows (SPSS Inc., Chicago, Illinois, USA ). P-verdi mindre enn 0,05 ble ansett som statistisk signifikant.
Resultater
Utbredelse av EBVaGC og tilknytning til klinisk-patologiske egenskaper
Åtti en prakteksemplarer av GC ble testet for EBV positivitet bruker EBER in situ
hybridisering. Frekvensen av EBVaGC var 14,8% (12 av 81) i hvilken det EBER positivt signal ble begrenset til kjernen av carcinoma-celler (figur 1). De EBVaGC prøvene har en variabel prosentandel av fargede tumorceller som viser mer enn 75% i løpet av seks tilfeller (Resultat: 3), mellom 50 og 75% i tre tilfeller og mindre enn 50% i de tre gjenværende tilfeller. De EBVaGC tilfellene var assosiert med alder ved diagnose (P
= 0,009, Tabell 2). Faktisk ble 9 av de 12 pasientene med EBVaGC alderen 45 til 60 år gammel. I tillegg ble en trend mot krets observert med tumor differensiering og venøs invasjon som den statisk betydelige verdier ikke ble nådd (P
= 0,07 og P =
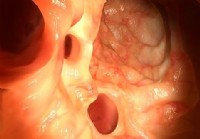
0,09 henholdsvis, tabell 2). Figur 1 Påvisning av Epstein-Barr virus kodet små RNA (Ebers) ved in-situ hybridisering i magekreft vev. A: H & E farging. B: Intens EBER positiv farging (immuno-stillingen 3+) i kjernen av tumorceller (opprinnelig forstørrelse x 40). C: Moderat EBER positiv farging (immuno-stillingen 2+) i kjernen av tumorceller (opprinnelig forstørrelse x 40). D: Svak EBER positiv farging (immuno-stillingen 1+) i kjernen av tumorceller (opprinnelig forstørrelse x 40). E: Positiv kontroll representeres ved en kjent Eber-positive NPC vev (opprinnelig forstørrelse x 40). F: magekarsinom vev negative for EBER (opprinnelig forstørrelse × 10)
Tabell 2 Sammenheng mellom EBVaGC og klinisk-patologisk parametre
Kliniske Kjennetegn
N
EBV uttrykk *
|
Negativ (%)
| Positiv (%)
Kjønn
Mann fra 48
42 (87,5)
6 (12,5)
Kvinne
33
27 (81,8)
6 (18.2)
p
-verdi
0,47
Age
< 45
17
17 (100)
0 (0)
45-60
30
21 (70)
9 (30)
> 60
34
31 (91,2)
3 (8,8)
p
-verdi
0,009
TNM
I-II
9
9 (100)
0 (0)
III-IV
47
40 (87,1)
7 (14,9)
p
-verdi
0,216
Anatomisk nettstedet
antrum
48
41 (85,4)
7 (14,6)
Body
22
19 (86,4)
3 ( 13.6)
Cardia
8
6 (75)
2 (25)
p
-verdi
0,72
Differensiering
Poor
43
36 (83,7)
7 (16,3)
Moderat
26
25 (96,2)
1 (3,8)
Vel
9
6 (66,7)
3 (33,3)
p
-verdi
0,075
Lauren typen
Tarm
47
38 (80,9)
9 (19.1)
Diffuse
34
31 (91,2)
3 (8,8)
p
-verdi
0,197
Tumor størrelse
< 5 cm
9
8 (88,9)
1 (11,1)
> 5 cm
65
54 (83,1)
11 (16,9)
p
-verdi
0,658
venøs invasjon
négatif
61
54 (88,5)
7 (11,5)
Positif
18
13 (72,2)
5 (27,8)
p
-verdi
0,09
* EBV-ekspresjon ble bestemt ved EBER in-situ-hybridisering. Negative tilfeller viste en score = 0 og positive tilfeller viste en score = 1+ til 3+
kvantitativ vurdering av EBV i GC
Q-PCR-analyse ble utført på 18 magekarsinom prøver.; blant dem 12 vises EBER positivitet begrenset til celler kjerner og 6 var negative.
viral DNA kopi nummer ble anslått av det absolutt kvantifisering metode med serie fortynning av rekombinant plasmid DNA (pGEMT /BamHI-W), som ekstern standard. Standardkurve ble etablert ved å plotte utgangs plasmid-kopitall mot terskelsyklusen (Ct), som viser en lineær kvantifisering over et område fra 10 6 til 10 kopier pr. Reaksjon
Representative eksempler er vist på figur 2 hvor tilfeller GC10 og GC3 (Ct = 15 og 22 henholdsvis) vil vise sterk EBER nukleær farging mens GC8 (Ct = 30) oppviser en svak EBER uttrykket (figur 2). Q-PCR data er i samsvar med EBER in situ
hybridisering resultat. Faktisk, de 6 prøvene med intens EBER farging (scorer 3+), viste høy EBV DNA kopiantall, mens tilfeller med svak EBER uttrykk (poengsum 1+) samsvarer med lav viral DNA-kopi (tabell 3). Figur 2 Fastsettelse av EBV DNA kopiere nummer av Q-PCR. Representative eksempler på GC tilfeller med høy (GC10), middels (GC3) og lav (GC8) EBV DNA kopier nummer. Den horisontale linjen markerer grensen som brukes for vurdering av Ct verdi.
Tabell 3 EBV DNA kopiantall og EBER flekker poengsum på 12 EBV positive prøver og 6 negative tilfeller
GC Cases
Histologisk
Skriv
Tumor FBT-Parts.com Site
Ct * product: (gjennomsnitt)
EBV DNA *
kopier /ug
EBER farging
scorer
10
Tarm
antrum
15
6,105
3+
3
Tarm
antrum
22
4,103
3+
12
Tarm
antrum
23
103
3+
9
Diffuse
antrum
22,8
103
3+
2
Tarm
Body
25
5,102
3+
4
Tarm
antrum
26
102
3+
6
Tarm
Proximal
27
8,101
2+
7
Diffuse
Body
28
5,101
2+
11
Tarm
Proximal
28
5,101
2+
1 Tarm
antrum
30
< 10
1+
5
Tarm
antrum
30
< 10
1+
8
Diffuse
Body
30
< 10
1+
13
Diffuse
antrum
ND
ND
0
14
Diffuse
Body
ND
ND
0
15
Diffuse
antrum
ND
ND
0
16
Diffuse
antrum
ND
ND
0
17
Tarm
antrum
ND
ND
0
18
Tarm
Body
ND
ND
0 product: * EBV DNA kopiantall ble anslått av Q-PCR der absolutt kvantifisering metoden. Ct: terskel Cycle. ND: Ikke Bestemt
Genotyping av EBVaGC stammer
å undersøke videre EBVaGC i våre 12 tunisiske eksemplarer, har vi undersøkt den genetiske profilen til EBV stammer bestemme forskjellige typer og varianter. En unik profil i alle undersøkte prøver er observert (figur 3). Det var preget av A /D-typer, prototype F, oppbevaring av Xho
I restriksjonssete og 30 bp del-lmp1 variant. Vi merker oss at A og B typer forme to ulike EBV stammer ble funnet i ett tilfelle av EBVaGC prøver (figur 3). Figur 3 Genotyping av EBVaGC stammer. A: PCR -RFLP av BamHI-F region som viser f variant (tilstedeværelse av to bånd med 128 bp og 71 bp) for styring C666-1 cellelinje (kolonne 1) og ett bånd på 199 bp for F-varianten for tilfeller GC2 til GC6 (spor 2-6). B: PCR-RFLP av BamHI-W1 /I1 region. DNA-fragmenter ble fordøyd med BamHI
gir to band av 139 og 67 bp (type D) for GC2 til GC6 (spor 2-6) eller ett band på 245 bp (type C) for kontroll C666-1 cellelinje ( spor 1). C: EBNA3C regionen. EBV stammer av type A og B tilsvarer DNA-fragment på 153 og 246 bp respektivt. Sak GC6 viser dual infeksjon med både type A og B virus. B95.8 er en type A virus som ble brukt som en kontroll (kolonne 1). D: Xhol
polymorfisme i exon 1 i BNLF1 genet. PCR-produktet ble fordøyd av XhoI
å gi to band av 343 bp og 154 bp i XhoI
+ variant for GC2 å GC6 (spor 2-6). C666-1-cellelinjen er en positiv taps-Xhol
variant (kolonne 1).
EBV DNA-analyse ble også utført ved partiell sekvensering på det tredje ekson av BNLF1 genet som koder for aminosyrene 328-376 av lmp1 protein. For å bekrefte den 30 bp som koder for sletting aminosyrer 343-352 av lmp1 protein, sekvenssammenstilling viste syv aminosyrer endringer i forhold til den referansestamme B95.8. Det ble stadig funnet i kodon Q334R, L338S og S366A i alle EBVaGC prøver. De resterende aminosyrer erstatninger, men ble spesielt funnet i en eller to EBVaGC eksemplarer: H358L og L359V (GC2), P360H (GC9-10) og G365R (GC6). Når det gjelder våre tidligere data om tunisiske NPC prøver, fant vi tilsvarende resultater om de tre konstant aminosyrer erstatninger. I tillegg er de EBV-stammer som bærer S366A substitusjon vises mer knyttet til Tunisian prøvestykker i forhold til de som er definert i Kina (figur 4). Figur 4 Sammenligning av utledede aminosyresekvenser fra Tunisia GC med B95.8 prototype og tidligere publiserte lmp1 sekvenser: Cao: NPC cellelinje fra Shangai [27]; C666-1 (GenBank tilgangsnummer ABV54173) NPC 10 fra Hong Kong [28] og tunisiske NPC eksemplarer: CV4, CV5, CV6 [29]. Symboler (---) indikerer aminosyre sletting.
Hos friske EBV bærere, polymorfisme analyse viste også A- og D-typer, prototype F og XhoI + varianten, som ligner identifisere i EBVaGC prøver (data ikke vist).
Diskusjon
I denne studien ble 81 tilfeller av GC fra pasienter i sør-regionen i Tunisia undersøkt for tilstedeværelse av EBV og utbredelsen av EBVaGC var 14,8% (12 av 81). Det var kjent at forekomsten av EBVaGC varierer sterkt fra en geografisk område til et annet, og den høyeste frekvensen ble registrert i Tyskland (18%), mens den laveste en (3,9%) ble detektert i Peru [13-20, 31].
EBER in situ hybridisering
var den mest pålitelige metode som er rapportert i litteraturen for å detektere latente EBV i GC. Faktisk ble alle de overnevnte studiene utført etter denne metoden, inkludert vår studie. De variable mengdeforhold fra kjernetumorceller som uttrykker EBER viser i vår studie ble også utledes ved Truong et al., Som foreslår å være relatert med EBV-infeksjon forekommer i onkogene fremgangs EBVaGC [32]. Imidlertid må ytterligere undersøkelser utføres for å klargjøre denne observasjonen.
I tillegg viste vi at de heterogene data Eber uttrykket kan være korrelert med antallet av EBV DNA-kopier som anslått ved Q-PCR som støtter at denne metoden er pålitelig så rapportert tidligere [33].
med hensyn til klinisk-patologiske trekk ved EBVaGC, sterk sammenheng med alder ved diagnose ble observert (P
= 0,009, Tabell 3). Faktisk ble EBVaGC oftere funnet i gruppen av pasienter i alderen mellom 45 og 60 år gammel, noe som er i tråd med tidligere studier [19, 34]. Ingen annen statistisk sammenheng ble funnet, bortsett fra en tendens med tumor differensiering og venøs invasjon (P
= 0,075 og P
= 0,09 henholdsvis). Tidligere rapporter har indikert at EBVaGC var hyppig i mannlig, proksimale magen og svulster i diffuse typen [9, 14, 16, 35-39]. Nylig, i en stor meta-analyse, Carmago et al., Bekrefter disse foreningene i tillegg til alder ved diagnose [38]. I vår studie fant vi ikke forskjell i fordelingen av EBV i mannlige forhold til kvinnelige pasienter, i proksimale vs distal mage og diffuse vs intestinal histotype. Disse variasjonene mellom data kan forklares av bidrag av lokale risikofaktorer i patogenesen av EBV og også av størrelsen og charasteristics av kohorten.
Polymorphism analyse av de 12 EBVaGC saker viser utelukkende av type D, prototype F, Xho
i-oppbevaring og 30 bp del-lmp1 variant. Når det gjelder polymorfisme av EBNA-3C-genet, har vi funnet en type i alle EBVaGC tilfeller og en kombinasjon av type A og B ble funnet i bare ett tilfelle. Disse funnene er i tråd med fersk undersøkelse gjennomført på fire EBVaGC saker fra den sentrale regionen av Tunisia [34].
Overvekt av type A og prototype F ble også vist i tidligere rapporter uavhengig av geografisk opprinnelse EBVaGC som i Sør- Kina [35], sørlige Japan [40], og latinamerikanske land [41]. Interessant, variant-f ble spesielt beskrevet i NPC forbundet EBV stammer av det sørlige Asia [42]. I dette området ble det en overvekt av C-type og tap-XhoI
variant observert hos pasienter med EBVaGC eller NPC [35, 40], i motsetning til vår nåværende funn og tidligere data på tunisiske NPC pasienter [29, 43].
for bedre å definere genotype av EBVaGC stammer og sammenligne dem med de som er forbundet med NPC, vi gjennomført delvis sekvensering av ekson 3 av BNLF1 genet og sammenlignet dem med prototypen B95-8 og andre publiserte sekvenser fra asiatiske og tunisiske NPC EBV stammer [29, 43]. Den Q334R ble L338S og L338P finnes i alle våre EBV isolerer og var allerede rapportert i EBV stammer knyttet til NPC som stammer fra Kina eller Tunisia [29, 44]. Imidlertid er S366A spesifikke EBV isolater assosiert med tunisisk NPC og GC .. Ja, og basert på den forrige rapporten fra Edwards et al. [44] beskriver syv fylogenetisk forskjellige stammer av LMP1, kan vi foreslå at tunisiske isolater utgjøre en supplerende gruppe med en bestemt signatur (T366A), men denne hypotesen trenger bekreftelse ved sekvensering av hele BNLF1 genet i disse isolatene. Ifølge denne datoen, foreslår vi at polymorfisme i BNLF1 genet utgjør en ekstra argument på linje med de andre EBV polymorfismer (D-type, prototype F og XhoI +) støtter ulikheten av EBV stammer mellom de to geografiske regioner.
På derimot har vi beskrevet tidligere
andre del-lmp1 varianter (69 bp og 81 bp sletting spenner kodoner 334-353 og 345-371, henholdsvis) i tunisiske NPC pasienter [29, 43]. Disse variantene ble ikke funnet, og bare den 30 bp del-lmp1 varianten ble identifisert i alle tilfeller EBVaGC som allerede rapportert av Chen et al., I en stor studie utført på kinesiske GC-pasienter [35].
Vår polymorfisme analyse av EBV isolater i friske bærere åpenbart en identisk genotype til de av EBVaGC og NPC som tyder på at vanlige stammer er geografisk fordelt, men ikke i forbindelse med en spesifikk malignitet. Faktisk kunne utviklingen av EBV assosiert malignitet være korrelert til variasjon i potensiell immun anerkjennelse i ulike populasjoner og enkeltpersoner, som foreslått av Edwards et al, [44].
Konklusjon
utbredelsen av EBVaGC hos pasienter fra Sør Tunisia er 14,8% som er innenfor rekkevidde med innrapporterte data. Den EBVaGC ble hovedsakelig funnet i gruppen av pasienter i alderen fra 45 til 60 år gammel, og at EBV DNA nivå reflektere EBER status i magekreft.
Videre EBV stammer knyttet til tunisiske pasienter med GC eller NPC dele noen likheter som tyder på at sannsynligvis den samme EBV-stammen er forbundet med begge tumorer. Til sammen våre funn støtter ulike geografiske fordelingen av EBV stammer, men ikke deres begrensning til en tilhørende malignitet.
Erklæringer
Takk
Dette arbeidet ble støttet av en bevilgning på den tunisiske Ministry of høy utdanning og forskning . Vi ønsker å takke Mr M Jaoua for sekvense fasiliteter.
Forfatternes opprinnelige innsendte filer for Images Nedenfor er linkene til forfatternes opprinnelige innsendte filer for bilder. 12985_2011_1593_MOESM1_ESM.tiff Forfatteroriginalfilen for figur 1 12985_2011_1593_MOESM2_ESM.tiff Forfatteroriginalfilen for figur 2 12985_2011_1593_MOESM3_ESM.tiff Forfatteroriginalfilen for figur 3 12985_2011_1593_MOESM4_ESM.tiff Forfatteroriginalfilen for figur 4 Konkurrerende interesser
Forfatterne erklærer at de har ingen konkurrerende interesser.
|  Årsaker til magesmerter
Hva forårsaker magesmerter? Magesmerter er smerter eller ubehag som en person føler hvor som helst mellom den nedre delen av brystet til den distale lysken. Noen helsepersonell inkluderer mellomgulve
Årsaker til magesmerter
Hva forårsaker magesmerter? Magesmerter er smerter eller ubehag som en person føler hvor som helst mellom den nedre delen av brystet til den distale lysken. Noen helsepersonell inkluderer mellomgulve
 Hva forårsaker Steatorrhea?
Steatoré, eller fet avføring, oppstår når det er for mye fett som fordøyelsessystemet ikke kan absorbere. Lær om 15 vanlige årsaker til steatoré Steatoré, eller fet avføring, oppstår når det er for m
Hva forårsaker Steatorrhea?
Steatoré, eller fet avføring, oppstår når det er for mye fett som fordøyelsessystemet ikke kan absorbere. Lær om 15 vanlige årsaker til steatoré Steatoré, eller fet avføring, oppstår når det er for m
 Fordøyelsessykdommer:årsaker til at magen din gjør vondt
Maveproblemer Magen til alle kommer litt ut av slag fra tid til annen. Men i noen tilfeller, avhengig av symptomene dine, kan det hende du må oppsøke legen din. Gastritt Væsken som hjelper deg med
Fordøyelsessykdommer:årsaker til at magen din gjør vondt
Maveproblemer Magen til alle kommer litt ut av slag fra tid til annen. Men i noen tilfeller, avhengig av symptomene dine, kan det hende du må oppsøke legen din. Gastritt Væsken som hjelper deg med