Tumor suppressor MIR-24 begrenser magekreft progresjon av downregulating RegIV
Abstract
Bakgrunn
microRNAs er små kodende RNA som modulerer en rekke cellulære prosesser ved å regulere flere mål, som kan fremme eller hemme utvikling av ondartet atferd . Samlende tyder MIR-24 spiller viktige roller i menneskelige kreftutvikling. Imidlertid må det presise biologiske rolle i stor grad unnvikende. Denne studien undersøkte rollen MIR-24 i magekreft (GC).
Metoder
uttrykk for MIR-24 i GC vev sammenlignet med matchede ikke-tumorvev og GC-celler ble oppdaget av QRT-PCR. Syntetisk korte enkle eller doble RNA oligonukleotider og lentiviral vektorer ble brukt til å regulere MIR-24 uttrykk i GC celler for å undersøke dens funksjon in vitro Hotell og in vivo
.
Resultater
MIR-24 var signifikant nedregulert i GC vev sammenlignet med matchede ikke-tumorvev, og er forbundet med tumor differensiering. Ektopisk uttrykk for MIR-24 i SGC-7901 GC celler trykt celleproliferasjon, migrasjon og invasjon in vitro
samt tumorigenicity in vivo
ved å indusere cellesyklus arrest i G0 /G1 fase og fremme celle apoptose. Videre identifiserte vi RegIV som et mål av MIR-24 og demonstrert at Mir-24 regulert RegIV uttrykk via binding sin 3 'uoversatt region.
Konklusjoner
Mir-24 fungerer som en roman tumor suppressor i GC og anti -oncogenic virksomheten kan medføre sin hemming av målgenet RegIV. Disse funnene antyder muligheten for MIR-24 som et terapeutisk mål i GC.
Nøkkelord
MIR-24 RegIV Magekreft Proliferation Invasion metastaser Bakgrunn
Magekreft (GC) er en av de vanligste kreft hos mennesker. Selv om forekomst og dødelighet har gått ned over hele verden i løpet av de siste 20 årene, GC fortsatt rangert som den fjerde mest vanlige og den nest mest dødelige kreft på verdensbasis [1]. Selv om forskning i GC har gjort store fremskritt, de molekylære mekanismene bak GC fortsatt ikke fullstendig klarlagt.
MicroRNAs (mirnas) er små kodende, dobbel-strandet RNA-molekyler som kan modulere uttrykket av målgener ved å påvirke post- transkripsjons prosesser som regulerer genuttrykk. mirnas gjenkjenne mål-mRNA basert på fullstendig eller ufullstendig sekvens komplementaritet og hindre produksjon av proteinet ved å binde seg til det 3'-ikke-translaterte region (UTR) eller den åpne leseramme (ORF) av mål-mRNA [2, 3]. mirnas har blitt vist å fungere i cellulær proliferasjon og differensieringsprosesser, inklusive den epiteliale-mesenkymale overgang [4] og endodermal differensiering av humane embryoniske stamceller [5]. I tillegg til de generelle cellulære funksjoner av miRNA, disse molekylene spiller også en viktig rolle i en rekke humane sykdommer. Flere studier har vist at misregulation av mirnas har sammenheng med kreft [6, 7], hvori de fungerer enten som onkogener eller tumor-suppressorer. mirnas har blitt rapportert å være involvert i akutt myeloid leukemi, brystcancer, ikke-småcellet lungekarsinom, leverkreft, tykktarmskreft, GC og andre kreftsykdommer [8-13]. Overekspresjon eller nedregulering av miRNAs fører til spredning av protein uttrykk, derfor bidra til tumorigenesis ved å modulere spredning, angiogenese og invasjon. Forskerne fant at miRNA ekspresjonsprofiler kan brukes til å klassifisere humane kreftformer, og dette funn var før anvendelsen av messenger RNA-profiler for klassifisering av dårlig differensierte tumorer [14, 15].
Aberrant ekspresjon av spesifikke mirnas i GC har vært tidligere rapportert og ble korrelert med utviklingen og prognose for GC [16-18]. Vi har tidligere funnet at MIR-24, som kan tjene som en tumor suppressor gjennom et målområde polymorfisme [19], ble en signifikant nedregulert miRNA i GC-celler og vev, sammenlignet med ikke-tumorvev. I denne studien har vi analysert MIR-24 uttrykk nivåer i GC vev sammenlignet med matchede ikke-tumorvev, og vurderes korrelasjoner mellom MIR-24 nivå og clinicopathologic parametere. Vi evaluerte påvirkningen av overekspresjon av MIR-24 på vekst og apoptose av SGC-7901 GC celler både in vitro Hotell og in vivo
. Vi har også bestemt den biologiske effekten av nedregulering av MIR-24 uttrykk på GC-celler. Videre identifiserte vi RegIV (regenererende holme-avledet familie, medlem 4) som en av målgener av MIR-24. Vi hypotese at Mir-24 kan regulere invasjon og metastasering av GC-celler ved direkte rettet mot RegIV genet.
Resultater
Overuttrykte MIR-24 hemmer GC celleproliferasjon
å utforske uttrykk for MIR-24 i GC, kvantitativ sanntids RT-PCR (QRT-PCR) ble utført. Uttrykk av MIR-24 ble generelt nedregulert i ni GC cellelinjer i forhold til de udødeliggjort normale mageslimhinne epiteliale celle GES-en (figur 1A). Uttrykk av MIR-24 ble undersøkt videre med QRT-PCR i tumorvev og matchet ikke-tumorvev fra 63 GC pasienter (Figur 1b). Den gjennomsnittlige uttrykk nivået av MIR-24 var signifikant nedregulert i tumorvev sammenlignet med matchede ikke-tumorvev (figur 1C). Sammen utgjør disse resultatene gitt sterke bevis for at Mir-24 var signifikant nedregulert i GC. Figur 1 MIR-24 er betydelig nedregulert i GC og hemmet SGC-7901 cellevekst. (A) Relativ uttrykk for MIR-24 i ni GC cellelinjer sammenlignet med GES-1 bestemt ved QRT-PCR av tre uavhengige eksperimenter (** P < 0,01, *** P < 0,001). (B) Relativ uttrykk for MIR-24 i kirurgiske prøver av 63 GC bestemmes av QRT-PCR. Alle forsøk ble utført in triplo. Data er vist som -ΔΔCT verdier, S.D. ikke vist. (C) Et gjennomsnitt og standardavvik av MIR-24 ekspresjonsnivåer i kirurgiske prøver av 63 GC vev og passet ikke-tumorvev er vist. Data er presentert som to-ΔCT verdier (** P < 0,05). U6 snRNA ble anvendt for normalisering. (D) MIR-24 hemmet veksten av SGC-7901-celler som detektert ved WST, mens anti-MIR-24 øket cellevekst (* P < 0,05). Data er vist som gjennomsnitt ± S.D. av tre uavhengige eksperimenter.
Vi neste undersøkt sammenhengen mellom uttrykksnivået MIR-24 og clinicopathologic faktorer i menneskelig GC. De kliniske og patologiske kjennetegn ved 63 GC pasienter er gitt i tabell 1. Basert på relative uttrykk prosenter av MIR-24 /U6 = 1, ble sakene delt inn i to grupper: Mir-24 high-uttrykk gruppe (n = 29) og Mir-24 lav-uttrykk gruppe (n = 34). Mir-24 lav-uttrykk gruppen viste signifikant lavere tumor differensiering (P = 0,021). Men den MIR-24 uttrykk nivå ikke viser noen sammenheng med alder, kjønn, tumorlokalisering, dybden av lokal utbredelse, lymfeknutemetastaser, eller TNM stage.Table 1 Forholdet mellom MIR-24 uttrykksnivå og clinicopathologic variabler i 63 GC pasienter
Clinicopathologic parametere Book MIR-24 uttrykk
P-verdi
Høy (n = 29)
lav (n = 34)
Kjønn
Mann fra 17
21
0,799
Kvinne
12
13
Alder (år)
< 60
15
18
0,923
≥60
14
16
Tumor plassering
Distal tredje
8
8
0.310
Middle tredje
6
13
Proximal tredje
15
13
WHO klassifisering
Adenocarcinoma 25
28
0,155
Signet-ring cellekreft
1 5
mucinous adenokarsinom
3
1 Differensiering
Vel, moderat
15
8
0,021
Dårlig
14
26
Borrmann klassifisering
I /II
11
12
0,828
III /IV
18
22
Lokal invasjon
T1 /T2
1 7
0,098
T3 /T4
28
27
lymfeknutemetastase
Ingen
6
4
0,535
Ja
23
30
Distant metastase
Ingen
28
31
0,724
Ja
1 3
TNM stadium
I /II
7
8
0,955
III /IV
22
26
å undersøke den biologiske funksjonen til MIR-24 i utvikling og progresjon av GC, transfektert vi SGC-7901 celler med MIR-24 etterligner (SGC-7901 /MIR-24) eller MIR-24 inhibitor (SGC-7901 /anti-MIR-24). Som vist i tilleggsfiler 1: Figur S1 A og S1B, valgte vi 100 nM som egnet konsentrasjon i etterfølgende eksperimenter. Ektopisk uttrykk for MIR-24 i SGC-7901-celler ble bekreftet ved QRT-PCR. Deretter undersøkte vi celleproliferasjon ved WST analysen. Overekspresjon av MIR-24 hemmet vekst på SGC-7901 celler sammenlignet med MIR-kontroll transfekterte celler (P < 0,05), mens anti-MIR-24 økt cellevekst aktiviteter (P < 0,05, figur 1D).
Overuttrykte MIR-24 hemmer migrasjon og invasjon av GC-celler
Neste vi evaluert effekten av MIR-24 på celle migrasjon og invasjon i GC. Celle migrasjon og invasjon analyser viste at overekspresjon av MIR-24 trykkes celle migrasjon (SGC-7901 /MIR-24 gruppen, 63,0 ± 3,6 celler per felt, kontrollgruppen, 126,3 ± 7,0 celler per felt; p < 0,05) og invasjon ( SGC-7901 /MIR-24 gruppen, 34,7 ± 2,1 celler per felt, kontrollgruppen, 63,7 ± 3,5 celler per felt; p < 0,05) (figur 2A, B). I kontrast, knockdown av MIR-24 betydelig økt celle migrasjon (SGC-7901 /anti-MIR-24 gruppen, 182,3 ± 5,0 celler per felt, kontrollgruppen, 105,3 ± 3,8 celler per felt; p < 0,01) og invasjon (SGC -7901 /anti-MIR-24 gruppen, 124,3 ± 3,8 celler per felt, kontrollgruppen, 62,7 ± 2,5 per felt; P < 0,01) (figur 2A, C). Figur 2 MIR-24 hemmet migrasjon og invasjon av SGC-7901 celler. (A) Representative felt av SGC-7901 celler i transwell analyser. (B, C) Representative felt av SGC-7901 celler i migrasjon og invasjon analyser, henholdsvis. Gjennomsnittlig antall migrasjon og invasjon celler per felt fra tre uavhengige forsøk (* P < 0,05, ** P < 0,01).
Overekspresjon av MIR-24 fremmer apoptose av GC-celler og induserer cellesyklus arrest i G0 /G1 fase
Gitt at observerte cellevekst kan påvirkes av satsene for apoptose og cellesyklusprogresjon, undersøkte vi effekten av MIR-24 på apoptose og cellesyklus i vivo
av flowcytometri. Vi fant at ca 12-15% av SGC-7901 /MIR-24 celler utstilt morfologiske egenskaper som er typisk for apoptose, inkludert kondensert kromatin og kjernekraft fragmentering av Hoechst33342 farging for DNA innhold. I kontrast, når man tar høyde for den sjeldne spontan apoptose i SGC-7901 celler, SGC-7901 /anti-MIR-24 gruppen viste ingen signifikante endringer ved Hoechst33342 farging (figur 3A). Flowcytometri viste at apoptotiske hastigheten øker betydelig hos SGC-7901 /MIR-24 celler sammenlignet med kontrollceller (13,62% ± 1,25% vs. 6,15% ± 0,95%, henholdsvis; P < 0,01), og betydelig redusert i SGC -7901 /anti-MIR-24 sammenlignet med kontroller (3,92% ± 0,52% vs. 6,35% ± 0,83%, henholdsvis; P < 0,05). (Figur 3B, D)
for ytterligere å belyse mekanismen av speil 24-formidlet vekstinhibering av GC-celler, ble cellesyklusanalyse utført (figur 3C, E). Ved oppregulering av MIR-24, prosentandelen av celler i G0 /G1 fase øket fra 40,51% ± 3,15% i kontrollene til 72,24% ± 3,65% (P 0,01), mens knockdown MIR-24 redusert prosentandelen av celler i G0 /G1 fasen fra 42,35% ± 2,78% hos kontrollene til 30,25% ± 1,25% (P < 0,05). Figur 3 MIR-24 indusert celle apoptose og G0 /G1 cellesyklus arrest. (A) Representative histogram som viser kjernefysisk morfologi SGC-7901 celler transient transfektert med 100 nM MIR-24 etterligner eller hemmer og deres respektive kontroller (Hoechst33342 flekker, opprinnelig forstørrelse 400 ×). (B) Representative histogrammet viser apoptose av SGC-7901 celler transient transfektert med 100 nM MIR-24 etterligner eller hemmer og deres respektive kontroller. (C) Representative histogram som viser cellesyklusen SGC-7901 celler transient transfektert med 100 nM MIR-24 etterligner eller hemmer og deres respektive kontroller. (D, E) Prosentandelen av apoptotiske og G0 /G1 faseceller av tre uavhengige eksperimenter, gjennomsnitt ± S.D. (* P < 0,05, ** P < 0,01).
RegIV er et mål gen av MIR-24
Til nærmere undersøke mekanismene for Mir-24 i GC, søkte vi etter kandidat målgener ved bioinformatikk. TargetScan, miRBase Tatget og Starbase ble brukt for å søke etter potensielle mål av MIR-24. Blant de antatte målene, ble RegIV identifisert som en av de målgener av MIR-24, og vi identifisert en potensiell MIR-24 bindingssetet i sin 3'UTR (Figur 4A). Deretter undersøkte vi uttrykket av RegIV i ni GC celler og GES-1. Vi fant at RegIV ble overuttrykt i ni GC-celler sammenlignet med GES-1, og oppviste en invers uttrykk mønster sammenlignet med MIR-24 (Tilleggs fil 2: Fig S2A og S2B). For å undersøke om 3'UTR av RegIV mRNA var et funksjonelt mål på MIR-24, ble luciferasereportergenet analyser utført. Vi evaluerte første aktiviteten av MIR-24 (eller MIR-kontroll) co-transfektert inn SGC-7901 celler med Luc-RegIV plasmid eller Luc-RegIV-mut plasmid (der den antatte MIR-24 bindingssetet ble mutert), sammen med PRL-TK-plasmid inneholdende det Renilla luciferasegenet som en intern kontroll (figur 4B). Celler ko-transfektert med MIR-24 viste en signifikant nedgang i luciferase-aktivitet sammenlignet med den MIR-kontrollgruppen (P 0,05). Imidlertid MIR-24 ko-transfektert med Luc-RegIV-mut plasmid viste ingen signifikant forskjell i reporter aktivitet sammenlignet med celler ko-transfektert med MIR-kontroll. Likeledes økte anti-MIR-24 luciferase-aktivitet av villtype-Luc-RegIV, men hadde ingen virkning på Luc-RegIV-mut plasmid (figur 4C; P < 0,05). Vi utførte også luciferasereportergenet assays i SNU-16 for å minimalisere påvirkning av endogene MIR-24. I begynnelsen ble ektopisk uttrykk for MIR-24 i SNU-16 celler bekreftet ved QRT-PCR. Ekspresjon av MIR-24 transfektert med MIR-24 etterligner var omtrent 50 ganger høyere enn den til mir-kontroll-gruppen i SNU-16 (P < 0,01), mens ingen statistisk forskjell med transfeksjon av anti-MIR-24 (tilleggsfiler 3: Figur S3A). Celler co-transfektert med MIR-24 viste en betydelig reduksjon av luciferase aktivitet sammenlignet med MIR-kontrollgruppen (P < 0,001), mens ingen statistisk forskjell med anti-MIR-24 transfeksjon (tilleggsfiler 3: Figur S3b og S3C) . Disse resultatene sterkt indikerte at 3'UTR av RegIV inneholder direkte bindingssteder for Mir-24. Figur 4 MIR-24 målrettet 3'UTR av RegIV genet og farging av RegIV i mage vev. (A) Skjematisk graf av de antatte bindingsseter fra Mir-24 i RegIV 3'UTR. RegIV-mut indikerer RegIV-3'UTR med mutasjon i MIR-24-bindingssteder. (B) MIR-24 ligner downregulated aktiviteten til en luciferase reporter som inneholder vill-type RegIV 3'UTR (* P < 0,05), men ikke reporteren med mutant RegIV 3'UTR. (C) Anti-MIR-24 økte luciferase aktivitet av villtype-Luc-RegIV (* P < 0,05), men hadde ingen effekt på den mutante. (D) Førti-åtte timer etter MIR-24 etterligne og kontroll transfeksjon av SGC-7901 celler, protein nivå av RegIV ble betydelig redusert sammenlignet med MIR-kontroll som bestemmes av ELISA. Anti-MIR-24 økte ekspresjon av RegIV sammenlignet med anti-MIR-kontroll (* P 0,05, ** P 0,01). (E) Tjuefire timer etter MIR-24 etterligne eller inhibitor og den respektive styre transfeksjon i SGC-7901-celler, var det ingen forskjell i mRNA-nivået av RegIV sammenlignet med MIR-kontroll som bestemt ved QRT-PCR (P >0,05). Data er vist som gjennomsnitt ± S.D. av tre uavhengige eksperimenter. (F) Ingen ekspresjon av RegIV ble detektert i normal gastrisk mucosa. (G) RegIV ble uttrykt i tarm-metaplasia av magen. (H) Sterk uttrykk for RegIV ble observert i mage signet-ring cell carcinoma.
Neste vi undersøkte MIR-24 regulering av RegIV mRNA og proteinnivåer i transfekterte SGC-7901 celler. ELISA-analyse viste at RegIV protein nivåer ble sterkt undertrykt i SGC-7901 /MIR-24 celler, mens RegIV protein nivåer ble oppregulert i SGC-7901 /anti-Mir-24 celler (figur 4D; * P < 0,05, ** P < 0,01). QRT-PCR-analyse viste ingen forskjell i nivået av RegIV mRNA i alle transfekterte cellegrupper (figur 4E; P > 0,05). Samtidig vi har oppdaget at ekspresjon av c-myc i GC-celler som positiv kontroll transfektert med MIR-24 [20]. Vi fant ut at Mir-24 downregulated RegIV og c-MYC (tilleggsfiler 4: Figur S4A og S4B).
Tidligere studier har vist at RegIV var forbundet med spredning og metastasering av GC. Vi neste brukte immunhistokjemisk analyse for å vurdere ekspresjonen av RegIV i GC. Våre funn bekreftet overekspresjon av RegIV i GC. Proteininnholdet av RegIV i normal gastrisk mucosa var lavere enn intestinal metaplasi av mage og gastrisk signet-ring cellecarcinom (figur 4F-H). For å vurdere den kliniske relevansen av disse funnene, undersøkte vi sammenhengen mellom uttrykk for RegIV og Mir-24 i GC vev. Vi fant at RegIV og MIR-24 utviste en invers uttrykk mønster i GC vev (tabell 2). Disse resultatene støtter tanken om at nedregulering av MIR-24 resulterte i økte proteinnivåer RegIV i GC.Table 2 RegIV og Mir-24 utstillings inverse uttrykk mønster i GC
MIR-24 uttrykk
P-verdi
lav
Høy
Reg IV (IHC)
Negativ
9
16
0,038 *
Positive
25
13 product: * P
< 0,05 ble ansett som statistisk signifikant
Overuttrykte MIR-24 hemmer tumorigenicity in vivo
Gitt at Mir-24 forbedret. spredning av GC celler in vitro
, undersøkte vi om Mir-24 kan påvirke tumorigenicity in vivo
. Retrovirus-mediert SGC-7901 /MIR-24 og SGC-7901 /MIR-kontroll stabile cellelinjer ble oppnådd som beskrevet i metodedelen. SGC-7901 /RV-MIR-24 og SGC-7901 /RV-MIR-kontrollceller ble injisert subkutant i fire uker gamle hann nakne mus, og tumordannelse ble overvåket. Svulster vokste saktere i SGC-7901 /RV-MIR-24 gruppen enn de i SGC-7901 /RV-MIR-kontrollgruppen (figur 5A). Den gjennomsnittlige tumorvolumet i mus inokulert med SGC-7901 /RV-MIR-24 celler på dag 28 var betydelig mindre i forhold til mus inokulert med SGC-7901 /RV-MIR-kontrollceller (397.45 ± 93,07 mm
3 vs. 1083.56 ± 101,56 mm 3, henholdsvis) (figur 5B og C, P < 0,05). Figur 5 MIR-24 hemmet tumorigenicity og spredning in vivo. (A) Fotografi av svulster avledet fra RV-MIR-24 eller RV-MIR-kontrollceller i nakne mus. (B) Vekstkinetikken for tumorer i nakne mus. Tumordiametere ble målt etter 7 dager. Volumet av de nakne mus indikert ved barer, S.D. (* P < 0,05). (C) Den gjennomsnittlige vekten av tumorer i nakne mus. Data er vist som gjennomsnitt ± S.D. (* P < 0,05). (D) Representative fotografier av immunohistokjemisk analyse av Ki-67-antigenet i tumorer i nakne mus (opprinnelig forstørrelse 200 x). (E) Representant proliferativ indeks av svulster i RV-MIR-24 og RV-MIR-kontroll nakne mus. Data er vist som gjennomsnitt ± S.D. (** P < 0,05). (F) Expression nivåer av RegIV i tumorvev av nakne mus. Data er vist som gjennomsnitt ± S.D. (* P < 0,05). (G) Expression nivåer av RegIV mRNA i tumorvev av nakne mus (P > 0,05). Data er vist som gjennomsnitt ± S.D. av tre uavhengige eksperimenter.
For å vurdere om tumorvekstinhibitering i SGC-7901 /RV-MIR-24 celler var delvis på grunn av undertrykkelse av spredning, analyser immunhistokjemiske av tumorvev ble utført. Som vist med Ki-67 antigen flekker, kan den reduserte tumorvekst i mus injisert med SGC-7901 /RV-MIR-24 celler være delvis på grunn av lavere spredning forårsaket av overekspresjon av MIR-24. Prosentandelen av Ki-67-antigen-positive celler var lavere i svulsten avledet fra SGC-7901 /RV-Mir-24 celler enn svulsten avledet fra SGC-7901 /RV-MIR-kontrollceller (37,1% ± 3,6% vs . 79,5% ± 5,2%, henholdsvis) (figur 5D og E; P < 0,01). Uttrykket nivå RegIV bestemt ved ELISA var lavere i svulsten avledet fra celler overekspresjon MIR-24 enn svulst avledet fra kontrollceller (1,77 ± 0,15 ng /ml i SGC-7901 /RV-MIR-24 vs 3,13 ± 0,25 ng /ml i SGC-7901 /RV-MIR-kontroll) (figur 5F; P < 0,05). Imidlertid var det ingen statistisk forskjell i den relative ekspresjon av mRNA RegIV (figur 5G). Derfor tumorigenitet av SGC-7901 /RV-MIR-24 celler ble betydelig redusert in vivo
.
Diskusjon
Samler bevis har støttet viktige roller for mirnas, som fungerer enten som tumor suppressors eller onkogener, i GC [21-23]. I tillegg har mange mirnas blitt vist å oppvise terapeutisk potensial. På grunn av deres funksjon som master-regulatorer av genomet og ny virkningsmekanisme, er mirnas ansett som en lovende teknologi for terapeutisk utvikling [24]. MIR-26a har antitumorigenic egenskaper og potensiell terapeutisk nytte for leverkreft in vitro og in vivo
, og er forbundet med rask og vedvarende inhibering av kreftcellevekst og meget spesifikk induksjon av tumor celledød [25]. Den spesifikke mekanismen av rollene til feilregulert mirnas i magekreft fortsatt ukjent.
I denne studien har vi demonstrert den hemmende effekten av MIR-24 på tumormetastase ved klinisk, cellulært og molekylært nivå og i en eksperimentell dyremodell. Vi følger detaljert mekanistisk eksperimentelle bevis for rollen som MIR-24 i GC ved å undertrykke uttrykket av RegIV. Undersøkelser viste at MIR-24 kan fungere som onkogenet i maligne effusjoner [26], munnhulekarsinomer [27, 28], prostatakreft [3] og lungekreft [29, 30], men kan virke som en tumor suppressor i tykktarmskreft [ ,,,0],19] og retinoblastomtumorer [31]. Lal A et al. fant at MIR-24 hemmet celledeling og cellesyklusprogresjon ved å undertrykke uttrykket av E2F2, MYC og andre cellesyklusregulerende gener ved å binde seg til "stenfrie" 3'UTR miRNA anerkjennelse elementer [20]. Wu J et al. rapporterte at transfeksjon av MIR-24 inn i GC-celler redusert ekspresjon av AE1-protein, noe som resulterte i inhibering av cellulær proliferasjon [32]. Men den komplette underliggende mekanismer for Mir-24 i GC er fortsatt ikke klart.
Vår rapport identifisert MIR-24 som kandidat tumor suppressor i GC. Vi har funnet nedregulering av MIR-24 ekspresjon i både GC vev og cellelinjer. Overekspresjon av MIR-24 i SGC-7901 GC cellene betydelig redusert spredning og invasjon både in vitro Hotell og in vivo
, avslører den potensielle terapeutiske effekten av MIR-24 i GC. Det motsatte resultat ble oppnådd når uttrykket av MIR-24 ble hemmet av anti-MIR-24. Sammen tyder disse resultater på at MIR-24 kan fungere som en tumor suppressor i human GC.
Mirnas bindes til perfekt eller imperfekt komplementære "seed" sekvenser i mål-mRNA, som fører til spalting av mål-mRNA eller inhibering av deres oversettelse [33] . I denne studien har vi identifisert RegIV som et mål gen av MIR-24. MIR-24 bundet med ufullstendig komplementaritet til RegIV mRNA, som resulterer i direkte translasjonell hemming av RegIV mRNA, men med ingen effekt på total mRNA stabilitet. Som de ulike handlingsmåter miRNA, gjorde MIR-24 ikke påvirke mRNA nivået av RegIV men translasjonell hemming. RegIV er et medlem av den humane Reg genfamilien, som deler sekvenslikhet med karbohydrat-bindende domene av C-type lektiner, og ble først isolert og karakterisert i human inflammatorisk tarmsykdom [34]. RegIV er en type av utskilte proteiner, og spiller en rolle biologisk avhengig av de ekstracellulære utskilte proteiner, slik at vi fant ELISA som en foretrukket metode for påvisning. Western blot ble også utført for å validere proteinnivået av RegIV, og vi oppnådd de tilsvarende resultater. Tidligere studier har rapportert at RegIV genet ble ofte overuttrykt i GC og er potensielt involvert i invasjonen, metastaser, og kreftutvikling av GC. RegIV uttrykk ble snevert begrenset i noncancerous vev [35, 36]. RegIV ekspresjon var markert høyere i pasienter med peritoneal metastaser sammenlignet med de uten peritoneal metastasering. RegIV kan akselerere peritoneal metastaser i GC og nivåer i lavage væske kan tjene som en god markør for peritoneal metastaser [37-39]. RegIV kan fungere som et serum biomarkør for GC pasienter og hemmer 5-FU-indusert apoptose ved induksjon av Bcl-2 og dihydropyrimidin dehydrogenase [40].
I denne studien identifiserte vi MIR-24 som en potensiell oppstrøms regulator av RegIV. Først overekspresjon av MIR-24 redusert aktiviteten til en luciferasereportergenet inneholder 3'UTR av RegIV, mens mutasjon av "seed region" områder i 3'UTR av RegIV avskaffet regulerende effekt av MIR-24. Det motsatte resultat ble oppnådd når vi brukte anti-MIR-24. For det andre humane vev GC uttrykt signifikant lavere nivåer av MIR-24 enn ikke-tumorvev, mens GC vev inneholder betydelig høyere nivåer av RegIV protein enn ikke-tumorvev. Tredje, overekspresjon av MIR-24 downregulated RegIV på proteinnivå, mens nedregulering av MIR-24 økte RegIV protein nivå. Fjerde, overekspresjon av MIR-24 var signifikant relatert til spredning og metastasering, noe som indikerer en funksjonell overlapping med RegIV. Alle disse resultatene tyder på at RegIV kan være et direkte mål gen av MIR-24 i GC.
Carcinogenesis er en serie påfølgende hendelser, inkludert løsgjøring, migrering, lokal invasjon, angiogenese, intravasation, overlevelse i sirkulasjonssystemet, og ekstravasasjon gjenvekst i forskjellige organer [41, 42]. Her viste vi at Mir-24 kunne regulere carcinogenesen av GC gjennom moduler spredning, migrasjon og lokal invasjon. Videre antyder våre bevis muligheten for MIR-24 som et terapeutisk mål i GC. Videre studier er nødvendig for å fullt ut forstå de detaljerte mekanismer for Mir-24 i GC kreftutvikling og som en potensiell terapeutisk tilnærming.
Metoder
Etisk uttalelse
Skriftlig informert samtykke ble innhentet fra alle deltakerne, og studieprotokoll ble godkjent av etiske komité for Ruijin Hospital, Shanghai Jiaotong University School of Medicine. Alle mus eksperimenter ble godkjent av Animal Care (PermitÉ20810) og bruk komité og utført i samsvar med de offisielle anbefalingene fra omsorg og bruk forsøksdyr av Ruijin Hospital, Shanghai Jiaotong University School of Medicine.
Cellelinjer
De menneskelige GC cellelinjer SNU-1, SNU-16, NCI-N87, og KATO III ble kjøpt fra American Type Culture Collection (ATCC). Cellelinjene SGC-7901, MKN-28, BGC-823, MKN-45, og AGS ble kjøpt fra Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. Den foreviget normal mage mucosal epitel cellelinje (GES-1) var en gave fra professor Feng Bi (Huaxi Medical University, Chengdu, Sichuan-provinsen, Kina). Den embryoniske nyrecellelinje 293 T ble bevart i vårt laboratorium og opprettholdt i Dulbeccos modifiserte Eagles medium (DMEM) med 10% føtalt bovint serum (FBS) ved 37 ° C i en fuktet atmosfære med 5% CO 2. Alle GC-celler ble dyrket i RPMI 1640 supplementert med 10% FBS i en fuktet inkubator med 5% CO 2 ved 37 ° C.
Vevsprøver
primær kreftvev og parvise tilstøtende ikke-tumorvev ble samlet fra pasienter med GC gikk radikal gastrektomi ved Kirurgisk avdeling, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine. De tilstøtende normale vev ble oppnådd i løpet av 5 cm fra primærcancer. Vevsprøver ble umiddelbart hurtigfrosset i flytende nitrogen og lagret i et kjøleskap ved -80 ° C. Vev for immunhistokjemi ble fiksert i 4% paraformaldehyde og parafin-embedded. Clinicopathological data ble gjennomgått, og TNM staging klassifikasjon var basert på kriterier for amerikanske Joint Committee on Cancer (AJCC, 6. utgave). Alle prøver ble verifisert ved patologisk undersøkelse.
RNA isolering og QRT-PCR
Total RNA ble isolert fra vevsprøver og cellelinjer ved hjelp Trizol reagens (Invitrogen, Carlsbad, CA, USA) i henhold til produsentens instruksjoner. Uttrykket nivåer av miRNAs ble evaluert ved bruk av hårnål-itTM mirnas qPCR Kvantitering Kit (GenePharma, Shanghai, Kina) etter produsentens protokoll. U6 small nuclear RNA (RNU6B; GenePharma) ble anvendt for normalisering. Den relative uttrykk forholdet mellom MIR-24 i hver sammenkoblet svulst og ikke-svulstvev ble beregnet ved hjelp av to -ΔΔCT metode. Mir-24 uttrykk nivået ble definert som oppregulert når den relative uttrykk forholdet var > 1, og definert som nedregulert når den relative uttrykk ratio var mindre enn en. Alle forfattere lese og godkjent den endelige manuskriptet.
 Har du perfekt bæsj?
Visse samtaleemner er bare tabu. Du spør aldri en kvinne om hun er gravid - med mindre du er helt sikker på at hun venter. Og det er mange samtaler som du absolutt ikke diskuterer under en familiemidd
Har du perfekt bæsj?
Visse samtaleemner er bare tabu. Du spør aldri en kvinne om hun er gravid - med mindre du er helt sikker på at hun venter. Og det er mange samtaler som du absolutt ikke diskuterer under en familiemidd
 Fungerer eplecidereddik for IBS?
Med tanke på bummen om helsefordelene med eplecidereddik, spør folk om det er nyttig for irritabel tarm. Hjemmemedisiner som ACV har en tendens til ikke å få forskningsfinansiering slik farmasøytiske
Fungerer eplecidereddik for IBS?
Med tanke på bummen om helsefordelene med eplecidereddik, spør folk om det er nyttig for irritabel tarm. Hjemmemedisiner som ACV har en tendens til ikke å få forskningsfinansiering slik farmasøytiske
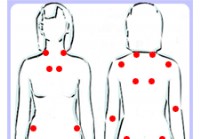 Fibromyalgi
Fakta du bør vite om fibromyalgi Illustrasjon av de ømme punktene ved fibromyalgi Fibromyalgi er et smertesyndrom preget av kronisk smerte, tretthet og ømhet ved berøring. Fibromyalgisyndrom er den v
Fibromyalgi
Fakta du bør vite om fibromyalgi Illustrasjon av de ømme punktene ved fibromyalgi Fibromyalgi er et smertesyndrom preget av kronisk smerte, tretthet og ømhet ved berøring. Fibromyalgisyndrom er den v