Overekspresjon av Snail er assosiert med lymfeknutemetastase og dårlig prognose for pasienter med magekreft
Abstract
Bakgrunn
Epitelial-mesenchymale overgang (EMT) spiller en betydelig rolle i tumorprogresjon og invasjon. Sneglen er en kjent regulator av EMT i forskjellige ondartede svulster. Denne studien undersøkte rollen Snail i magekreft.
Metoder
Vi undersøkte effekten av forstummet eller overuttrykt Snail hjelp lenti-viral konstruerer i magekreftceller. Immunhistokjemisk analyse av microarray fra 314 pasienter med adenokarsinom i ventrikkel (GC) ble benyttet for å bestemme snegle clinicopathological og prognostisk betydning. Differential genuttrykk i 45 GC prøver med Snail overekspresjon ble undersøkt ved hjelp av cDNA microarray analyse.
Resultater
stanse av Snail av shRNA redusert invasjon og migrasjon i GC cellelinjer. Motsatt Snail overekspresjon økt invasjon og migrasjon av magekreftceller, i takt med økt VEGF og MMP11. Snail overekspresjon (≥75% positive nukleær farging) ble også signifikant assosiert med tumorprogresjon (P
< 0,001), lymfeknutemetastaser (P
= 0,002), lymphovascular invasjon (P
= 0,002), og perinevral invasjon (P
= 0,002) i 314 GC pasienter, og med kortere overlevelse (P
= 0,023). cDNA microarray analyse avdekket 213 differensielt uttrykte gener i GC vev med Snail overekspresjon, inkludert gener relatert til metastasering og invasjon.
Konklusjon
Snail påvirker invasivitet /vandrende evne GCer betydelig, og kan også brukes som en prediktiv biomarkør for prognose eller aggressivitet GCer.
nøkkelord
magen Adenocarcinoma Snail lymfeknutemetastaser Survival Bakgrunn
epitelial-mesenchymale overgang (EMT), en utviklingsprosess der epitelceller redusere inter heft og skaffe myofibroblastic funksjoner, er kritisk til tumor progresjon [1-3]. Under EMT, oppstår betydelige endringer, inkludert nedregulering av epitel-markører som E-cadherin, translokasjon av β-catenin (dvs. dissosiasjon av membranøs β-catenin og translokasjon inn i kjernerommet), og oppregulering av mesenchymale markører så som vimentin og N -cadherin [3-6]. EMT er indusert av undertrykkelse av E-cadherin uttrykk av EMT regulatorer som sneglen, Slug, og Twist. The Snail familien av sink-finger transkripsjons repressors undertrykker direkte E-cadherin in vitro Hotell og in vivo
via et samspill mellom deres COOH-terminal regionen og 5
'- CACCTG-3 ' sekvens i E-cadherin promoter [7-9]. Snail er angivelig viktig i flere karsinomer, inkludert ikke-småcellet karsinom, eggstokkreft karsinom, uroteliale karsinom, og leverkreft [10-13]. Studier har også brukt immunhistokjemiske analyser for å vise den kliniske betydningen av Snail overekspresjon i adenokarsinom i ventrikkel (GC) [14, 15]. Imidlertid har noen rapporter om rollene til Snail i GC inkludert clinicopathological, prognostisk og funksjonell in vitro
analyser, samt genuttrykk resultater. Vi evaluerte derfor sneglen effekt på invasivitet /trekkende egenskaper i mage cancer cellelinjer, og også undersøkt muligheten for å sneglen blir brukt som en prediktiv markør for evaluering av dårlig prognose eller tumor aggressivitet i GC pasienter. Vi vurderte også genuttrykk mønster på 45 GC vev med Snail overekspresjon, ved hjelp av cDNA mikromatriser
. Metoder
shRNA lentivirus-mediert taushet overekspresjon av Snail i magekreftceller
menneskemagekreft cellelinjer SNU216 og SNU484 ble innhentet fra koreanske cellelinje Bank (KCLB) og ble godkjent av DNA profilering. Disse celler dyrket i RPMI1640 medium med 10% føtalt bovint serum (FBS), 100 U /ml penicillin og 100 ug /ml streptomycin (Hyclone, Ogden, Utah). Alle celler ble opprettholdt ved 37 ° C i 5% CO 2. Lentiviral-baserte RNA knockdown og overekspresjon ble brukt for å kneble og overekspresjon av Snail. Lentiviruses uttrykker enten ikke-target eller Snail
-targeted shRNAs ble brukt for å kneble; en PLKO lentiviral vektor rettet mot sneglen
eller en tom PLKO vektor ble brukt til overekspresjon av Snail i SNU216 og SNU484 celler. Lentivirus aksjer ble produsert ved hjelp av Virapower ™ lentiviral emballasje mix bruker 293ft cellelinje i henhold til produsentens protokoll (Invitrogen, Carlsbad, California). SNU216 og SNU484 celler dyrket til 50% konfluens ble inkubert i 24 timer i en 1: 1 fortynning av virus: media med 5 ug /ml polybrene. Etter en 24-timers restitusjonsperiode i komplett medium uten virus, ble polyklonale stabile cellelinjer valgt ut og opprettholdt i media inneholdende 5 ug /ml puromycin. Stanse eller overekspresjon av Snail ble bestemt ved RT-PCR og Western blotting.
Real time RT-PCR analyse av VEGF
, MMP11
, og Snail
i magekreftceller
Totalt cellulært RNA ble ekstrahert med TRIzol metode (Sigma-Aldrich, St. Louis, MO, USA). For RT-PCR-analyse, ble 2-ug porsjoner av RNA utsatt for cDNA-syntese med 200 U MMLV revers transkriptase og 0,5 pg oligo (dT) -15-primer (Promega, Madison, WI, USA). Kvantitativ real-time PCR ble utført med Rotor-Gene ™ System (Qiagen, Hilden, Tyskland) ved hjelp AccuPower 2 × Green qPCR Master Mix (Bioneer, Daejeon, Korea). cDNA i 1 ul av reaksjonsblandingen ble amplifisert med 0,5 U av GoTaq DNA-polymerase (Promega) og 10 pmol av hver av de følgende sense- og antisense-primere: GAPDH
5 '- TCCATGACAACTTTGGTATCG-3 ', 5 '- TGTAGCCAAATTCGTTGTCA-3 '; Snail
5 '- CTTCCTCTCCATACCTG-3 ', 5 '- CATAGTTAGTCACACCTCGT-3 '; VEGF
5 '- TTGCTGCTCTACCTCCACCA-3 ', 5 '- GCACACAGGATGGCTTGAA-3 '; MMP11
5 '- CTTGGCTGCTGTTGTGTGCT-3 ', 5-AGGTATGGAGCGATGTGACG-3 '. Den termiske sykluser profilen var: denaturering i 30 s ved 95 ° C, gløding i 30 s ved 52 ° C (avhengig av primerne anvendt), og forlengelse i 30 s ved 72 ° C. For semi-kvantitativ vurdering av ekspresjonsnivåer, ble 30 cykler benyttet for hver PCR-reaksjon. PCR-produktene ble størrelses-fraksjonert på 1,0% etidiumbromid /agarosegeler og kvantifisert under UV transilluminasjon. Terskelen syklus (CT) er definert som den fraksjonelle syklus nummeret som fluorescensen passerer en fast terskel over basislinjen. Relativ genekspresjon ble kvantifisert ved å benytte gjennomsnitts CT-verdien for hver tre eksemplarer prøve minus gjennomsnittlig tre eksemplarer CT verdi for GAPDH
. Forskjeller mellom kontrollen (tom vektor) og eksperimentgrupper (infisert med lentivirus) ble beregnet ved hjelp av formelen 2 - ([△ CT Lenti] - [△ CT kontroll]) og uttrykt som en fold endring i uttrykket i henhold til den komparative terskelsyklusen metode (2- △△ CT) [16].
Western blotting
Cellene ble høstet og avbrutt i lyseringsbuffer (1% Triton X-100, 1 mM EGTA, 1 mM EDTA, 10 mM Tris-HCl, pH 7,4 og proteaseinhibitorer). Celleavfall ble fjernet ved sentrifugering ved 10 000 x g
i 10 minutter ved 4 ° C. De resulterende supernatanter ble løst på en 12% SDS-PAGE under reduserende betingelser denaturert og overført til nitrocellulosemembraner. Membranene ble blokkert med 5% ikke-fett tørrmelk ved romtemperatur i 30 minutter og inkubert med primære antistoffer. Membranene ble vasket og inkubert med pepperrot-peroksidase-konjugert sekundært antistoff. Signalet ble visualisert ved hjelp av en forbedret kjemiluminescens (Amersham, Buckinghamshire, UK).
Cellemigrering og Matrigel invasjon assay
magekreft ble cellene høstet med 0,05% trypsin inneholdende 0,02% EDTA (Sigma-Aldrich), og suspendert i RPMI ved en konsentrasjon på 3 x 10 3 celler /brønn. Membranfilter (porestørrelse 8 pm) i engangs 96-brønners kjemotakse kamre (Neuro Probe, Gaithersburg, MD) ble forhåndsbelagt i 4 timer med 5 mg /ml fibronektin ved romtemperatur. Aliquoter (50 ul /brønn) av cellesuspensjonen ble lastet inn i de øvre kamrene, og 1% FBS ble lastet inn i det nedre kammer. Etter 24-h inkubasjon ble ikke-migrerende celler fjernet fra det øvre kammer med en bomullspinne; celler som er tilstede på den nedre overflate av innskuddet ble farget med Hoechst33342 (Sigma-Aldrich). Invaderende celler ble tellet under et fluorescens-mikroskop ved 10 x forstørrelse.
For Matrigel invasjon analysen, 3 x 104 celler /brønn ble sådd ut i det øvre kammer, som var belagt med Matrigel (5 mg /ml i kaldt medium, BD Transduction Laboratories, Franklin Lakes, NJ, USA), og serumfritt medium inneholdende 1% FBS eller kontroll kjøretøyet ble tilsatt til det nedre kammer. Etter 24 timers inkubasjon ble ikke-migrerende celler fjernet fra det øvre kammer med en bomullsdott, og celler som er tilstede på den nedre overflate av innskuddet ble farget med Hoechst33342 (Sigma-Aldrich). Invasive cellene ble deretter telles under et fluorescens mikroskop med 10x forstørrelse.
Tissue mikromatriser, immunhistokjemi, og tolkning av resultater, En semi-automatisert vev arrayer (Beecher Instruments, WI, USA) ble brukt til å konstruere microarray . Vi erholdt 3 vev kjerner, hver 0,6 mm i diameter, fra tumorvev tatt fra GC-pasienter. Kjerner ble ikke samlet inn fra mer invasive frontal eller sentrale områder av svulster. Objektglass ble oppvarmet ved 60 ° C i 30 min, deparaffinized med xylen, og deretter rehydrert. Seksjonene ble deretter neddykket i citrat-buffer antigen gjenfinning, mikrobølgebehandlet i antigen gjenfinning, behandlet med 3% hydrogenperoksyd i metanol for å slukke endogen peroksidase-aktivitet, og deretter inkubert med 1% bovint serumalbumin for å blokkere ikke-spesifikk binding. Deretter ble seksjonene inkubert med kanin-anti-sneglen (Abcam, UK) over natten ved 4 ° C. Normalt kaninserum ble anvendt som en negativ kontroll. Etter vasking, ble vevssnitt behandlet med sekundært antistoff, kontra med hematoxylin, dehydrert, og montert. Minst 500-tumorceller ble opptalt. Prosentandelen av celler med snegle + kjerner ble uttrykt i forhold til det totale antall tumorceller telles. Nuclear uttrykk for Snail ble gradert ved å klassifisere omfanget av positive nukleær farging som ≤50%, 50-75%, eller ≥75%.
Clinicopathological og overlevelse analyse av magekreft pasienter
Vi studerte en kohort av 314 GC pasienter som hver gjennomgikk en gastrostomi med lymfeknute disseksjon ved Pusan National University Hospital (PNUH) mellom 2005 og 2007. konsernet består 218 menn og 96 kvinner med en gjennomsnittsalder på 58,3 år (fra 25-83 år). Standard formalinfiksert og parafin-embedded seksjoner ble hentet fra Avdeling for patologi, PNUH, og National Biobank Korea, PNUH. Studien ble godkjent av Institutional Review Board. Ingen av pasientene fikk preoperativ strålebehandling og /eller kjemoterapi. Adjuvant kjemoterapi basert på 5-FU ble administrert på pasienter med trinn II, III og IV etter kurativ reseksjon. Vi vurderte flere clinicopathological faktorer i henhold til den koreanske Standardisert patologi rapport for magekreft, den japanske Klassifisering av magekarsinom (3 rd engelsk utgave), og den amerikanske Joint Committee on Cancer Staging Manual (7 th edition), inkludert tumorlokalisering, brutto utseende og størrelse, dybde av invasjonen, histologisk klassifisering (dvs. intestinal eller diffuse), og lymphovascular invasjonen [17-19]. Klinisk utfall for hver pasient ble fulgt fra dato for operasjonen til dødsdato eller 1. mars 2012. oppfølgingsperioder varierte fra ca 1 til 81,5 måneder (gjennomsnitt, 51,4 måneder). Tilfeller tapt for oppfølging eller død uavhengig av årsak annet enn magekreft ble sensurert fra overlevelse analyse. Clinicopathological funksjoner ble analysert ved hjelp av Student t
-test, den χ 2 test, eller Fishers eksakte test for å teste for forskjeller i Snail uttrykk. Akkumulert overlevelse tomter ble oppnådd ved bruk av Kaplan-Meier-metoden, og betydning ble sammenlignet med log-rank test. Prognostiske faktorer ble identifisert ved hjelp av Cox regresjon trinnvis metode (proporsjonal risikomodell), justert for pasientens alder, kjønn, tumorlokalisering, morfologisk type (intestinal versus diffuse). Statistisk signifikans ble satt til P
< 0,05. Statistiske beregninger ble utført med SPSS versjon 10.0 for Windows (SPSS Inc., Chicago, IL, USA).
CDNA microarray analyse av GC vev basert på sneglen overekspresjon
Totalt 45 friske GC vev ble hentet fra National biobank av Korea, PNUH, og CNUH; godkjenning er innhentet fra sine institusjonelle gjennomgang boards. Total RNA ble ekstrahert fra frisk-frosset vev ved hjelp av en Mirvana RNA Isolation Kit (Ambion Inc., Austin, TX). Fem hundre nanogram av total RNA ble benyttet for cDNA-syntese, etterfulgt av en forsterknings /merking trinn (in vitro transkripsjon
) ved hjelp av Illumina TotalPrep RNA Amplification Kit (Ambion) for å syntetisere biotin-merket cRNA. cRNA-konsentrasjoner ble målt ved hjelp av fremgangsmåten RiboGreen (Quant-iT RiboGreen RNA analysesett; Invitrogen-Molecular Probes, ON, Canada) ved hjelp av en Victor3 spektrofotometer (PerkinElmer, CT), og cRNA kvalitet ble bestemt på en 1% agarosegel. Merket, forsterket materiale (1500 ng per array) ble hybridisert til Illumina HumanHT-12 BeadChips v4.0, i henhold til produsentens instruksjoner (Illumina, San Diego, California). Array signalene ble utviklet av streptavidin-Cy3. Arrays ble skannet med en Illumina iscan system. Microarray data ble normalisert ved hjelp av quantile normaliseringsmetoden i Illumina BeadStudio programvare. Uttrykket nivået av hvert gen ble forvandlet til en log 2 base før videre analyse. Excel ble først og fremst brukt for statistiske analyser. Genekspresjon forskjeller ble ansett som statistisk signifikant dersom P
< 0,05; alle testene var 2-tailed. Cluster analyser ble utført ved hjelp av Cluster og Utforsker [20]. Genet ontologi (GO) program (http:... //David abcc ncifcrf gov /) ble brukt til å kategorisere gener inn i undergrupper basert på biologisk funksjon. Fishers eksakte test ble brukt for å avgjøre om proporsjonene av gener i hver kategori varierte med gruppen. GC vev ble videre delt inn i de med høyere (≥75%) og lavere (< 75%) nivåer av Snail uttrykk; differensial genekspresjon mellom gruppene ble identifisert. Primærmicroarray data er tilgjengelige i NCBI er GEO (Gene Expression Omnibus) database (http:.....? //Www NCBI NLM nih gov /geo /query /acc cgi acc = GSE38024).
Resultater
Regulering av migrasjon og invasjon av magekreftceller ved Snail
Lentiviral-basert RNA knockdown og overekspresjon tilnærminger ble benyttet for å bestemme Snail rolle i invasjonen og migrasjon av magekreft cellelinjer. SNU216 og SNU484 celler brukt i denne studien er etablert mage adenokarsinom cellelinjer fra koreanske pasienter. Disse cellene ble infisert med et lentivirus uttrykker enten ikke-target eller Snail
-targeted shRNAs for lyddemping. En PLKO lentiviral vektor som målrettet Snail
og en tom PLKO vektor ble brukt til å indusere Snail overekspresjon i SNU216 og SNU484 celler. Polyklonale stabile cellelinjer ble valgt ved hjelp puromycin. Snail
ble bestemt ved RT-PCR og Western blotting; stabil Snail
knockdown (sh-Snail) og Snail overekspresjon cellelinjer (OE-Snail) ble oppnådd (figur 1). Figur 1 Role of Snail i invasjonen og migrasjon av magekreft cellelinjer. A. SNU216 (øvre panel) og SNU484 (nedre panel) celler ble infisert med lentivirus uttrykker enten ikke-target shRNA (shNT
) eller Snail
shRNA på dag 0, og deretter høstet på dag 7 etter infeksjonen. Snail
knockdown ble bestemt ved RT-PCR og Western blotting; stabile cellelinjer ble generert for hver av cellelinjene (sh-sneglen). Stanse av Snail
i SNU216 og SNU484 celler indusert redusert migrasjon og invasjon. B. SNU216 (øvre panel) og SNU484 (nedre panel) celler ble infisert med lentivirus uttrykker enten en lentiviral PLKO vektor rettet mot sneglen
eller en tom PLKO vektor (EV) på dag 0, og deretter høstet på dag 7 etter infeksjon . Overekspresjon av sneglen ble bestemt ved RT-PCR og Western blotting; stabil cellelinje ble generert for hver av cellelinjene (O /E-snegle). Snail overekspresjon i SNU216 og SNU484 celler indusert økt migrasjon og invasjon. C. Snail overekspresjon indusert økt mRNA uttrykk for VEGF Hotell og MMP11
i SNU216 og SNU484 celler i real-time RT-PCR-analyse. Nedre panel viser representative RT-PCR tall for VEGF
, MMP11
, Snail
, og GAPDH
. Data viser gjennomsnitt ± SE av minst 3 uavhengige eksperimenter. * Viser P
< 0,05 av Student t
-test.
Å bestemme snegle roller i magekreft celle invasjon, målte vi kjemotaktisk invasjon av cellene ved hjelp av Transwell system med filtre pre-belagt med Matrigel. For å måle migrering av magekreftceller, analysert vi celle migrasjon ved hjelp av en Boyden kammer apparat. Stanse av Snail
av shRNA indusert redusert migrasjon og invasjon av SNU216 og SNU484 celler, som vist i figur 1A. I motsetning til den Snail
Silencing resultater, overekspresjon av Snail induserte økt migrasjon og invasjon av SNU216 og SNU484 celler, som vist i figur 1B. Overekspresjon av Snail var også assosiert med økt VEGF og MMP11 (figur 1C).
Effekt av Snail overekspresjon på tumor aggressivitet og GC pasient overlevelse
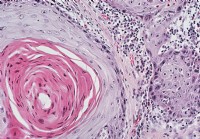
Positiv atom farging for sneglen på nivåer av ≤50%, 50-75%, og ≥75% ble observert i 13,4% (42/314), 52,2% (164/314), og 34,4% (108/314), henholdsvis, av de 314 pasientene i GC immunhistokjemisk analyse. Snail uttrykk ble notert i tarm og diffuse typen GCer (figur 2A, B). Snail overekspresjon (≥75% positivitet) signifikant korrelert med tumorstørrelse, brutto type, dybde av invasjonen, lymphovascular invasjon, perinevral invasjon, og lymfeknutemetastase (tabell 1). Snail overekspresjon var også assosiert med økt tumorstørrelse (P
= 0,028) og utgravd brutto type (P
< 0,001); og økt tumor invasivitet, dvs. høyere T trinn (P
< 0,001) og tilstedeværelsen av perinevral invasjon (P
< 0,001) og lymphovascular tumor emboli (P
= 0,002). Økt lymfeknutemetastase ble også knyttet til Snail overekspresjon (P
= 0,002) .I samsvar med ovennevnte data som viser positiv sammenheng mellom Snail overekspresjon og GC aggressivitet, Snail overekspresjon signifikant korrelert med total overlevelse blant GC pasienter (P
= 0,023) (figur 2C). En lineær sammenheng ble observert mellom økt kjernekraft uttrykk for sneglen og forkortet overlevelse (≤50%: 76,6 ± 2,7 måneder, 50-75%: 68,5 ± 2,0 måneder, ≥75%: 63,3 ± 2,8 måneder). Snail overekspresjon (≥75% positivitet) ble identifisert som en uavhengig prediktor for dårlig prognose i 314 pasienter med GC, justert for alder, kjønn, histologisk klassifisering, og svulsten plassering, ved hjelp av en Cox regresjon proporsjonal risikomodell (P
= 0,033; tabell 2). Figur 2 Snail uttrykk i mage adenokarsinom (GC) vevsprøver og Kaplan-Meir plott av total overlevelse av 314 GC pasienter. Snail ble stort sett uttrykt i kjernen av GC-celler (intestinal type (A), og diffus type (signetring) celler (B)) inkludert i vev array-prøver. Noen reaktive fibroblaster viste også Snail atom uttrykk (forstørrelse: × 400). C. Kaplan-Meier analyse av total overlevelse av GC pasienter basert på Snail uttrykk. En lineær sammenheng mellom økt Snail atom uttrykk og kortere overlevelse ble sett blant GC pasienter (P
= 0,023). Log-rank test ble brukt til å beregne P
verdier.
Tabell 1 Forholdet mellom Snail uttrykk og clinicopathological egenskaper hos 314 pasienter med magekreft
Antall pasienter (N = 314)
Snail Positivitet
p-verdi
< 75%
≥75%
Alder (år)
58,5 ± 10,6
59,1 ± 11,9
0,695
Sex
Mann fra 218
143
75
0,996
Kvinne
96
63
33
Tumor størrelse
≤4.0 cm
192
135
57
0,028
> 4,0 cm
122
71
51
beliggenhet
Øvre /Middle
167
112
55
0,561
Nedre
147
94
53
Invasion dybde
T1
160
127
33
< 0.001
T2
41
26
15
T3
68
33
35
T4
43
19
24
Gross typen
Forhøyet
77
51
26
< 0,001
Leilighet /deprimert
131
105
26
utgravde
106
50
56
Histologisk typen
Tarm
182
123
59
0,609
Diffuse
122
76
46
Blandet
10
7
3
perinevral invasjon
negativ
202
150
52
< 0.001
Positive
111
55
56
Lymphovascular emboli
Negativ
193
139
54
0,002
Positive
120
66
54
lymfeknutemetastaser
N0, N1
270
186
84
0,002
N2, N3
44
20
24
Tabell 2 Multivariat overlevelsesanalyse med Cox regresjonsmodell i 314 magekreftformer
variabler Book B
SE
HR (95% KI)
P
Age (≤59 versus > 59)
-0,438
0,264
0,645 (0,385 til 1,082)
0,097
Kjønn (mannlig versus kvinnelig)
-0,037
0,267
0,963 (0,571 til 1,626)
0,889 FBT-Parts.com Site (øvre og midtre versus lavere)
0,635
0,264
1,887 (1,126 -3,164)
0,016
Lauren (intestinal vs diffus)
-0,537
0,263
0,585 (0,349 til 0,978)
0,041
Snail (≥75% versus < 75 %)
-0,528
0,248
0,590 (0,363 til 0,958)
0.033
Merk: B, koeffisient; HR, hazard ratio; KI, konfidensintervall.
Identifikasjon av genuttrykksmønster basert på sneglen overekspresjon hjelp cDNA mikromatriser
cDNA mikromatriser ble brukt til å sammenligne genuttrykk profiler av 45 GC prøver. Vi identifiserte 213 gener som var forskjellig uttrykt ved betydelige nivåer (P
< 0,05) mellom GC prøver med høyere (≥75%) og lavere nivåer (< 75%) av Snail uttrykk (Tabell 3). Av disse 213 genene, 82 ble oppregulert og 131 ble nedregulert i GC prøvene med høyere nivåer (≥75%) av Snail uttrykk. Vi brukte hierarkisk clustering analyse for å vurdere de 213 genene og 45 GC eksemplarer; veiledet clustering analyse ga mønstre for prøver med høyere og lavere nivåer av Snail uttrykk gruppert i 2 forskjellige grupper, med unntak av én prøve med høyere nivåer av Snail uttrykk (figur 3). For å undersøke de biologiske funksjoner som er involvert i skjønns gener, utførte vi en GO kategorianalyse. Elleve gener ble assosiert med regulering kreftcelle-ECM adhesjon (P
< 0,021) og ECM protein regulering (P
< 0,028, Tabell 4). De fleste har vært innblandet i kreft. ONECUT1
, ADAMTS
, IFNAR2
, MSR1
, og SORL1
påvirke migrasjon eller metastasering, en prosess som involverer festing av kreftceller til basalmembran, nedbrytning av lokale bindevev, og penetrasjon og migrasjon av tumorceller gjennom stroma [21-25] .table 3 Gener forskjellig uttrykt i GC prøver med høyere nivåer av Snail uttrykk
PROBE_ID
SYMBOL
NAVN
Gener oppregulert i prøver med høyere nivåer (≥75%) av Snail uttrykk (P
< 0,05)
ILMN_2374449
SPP1
Utskilt phosphoprotein en
ILMN_2337923
TPD52L1
Tumor protein D52-lignende en
ILMN_1679838
WBP5
WW domene bindende protein 5
ILMN_2078592
C6orf105
androgen-avhengige TFPI regulerende protein
ILMN_1714383
TPD52L1
Tumor protein D52-lignende en
ILMN_1674817
C1orf115
kromosom 1 åpen leseramme 115
ILMN_1813561
Scin
Scinderin
ILMN_1759818
SORL1
Sortilin-relaterte reseptor, L (DLR klasse) A gjentar inneholder
ILMN_1745686
MFHAS1
Ondartet fibrøst histiocytoma forsterket sekvens en
ILMN_2060115
SORL1
Sortilin relaterte reseptor, L (DLR klasse) A gjentar inneholder
ILMN_2337263
PKIB
Protein kinase (cAMP -avhengig, katalytisk) hemmer beta
ILMN_2173835
FTHL3
Ferritin, tung polypeptid en pseudogen 3
ILMN_1791057
IFNAR2
Interferon (alfa, beta og omega) reseptor 2
ILMN_1807114
LOC255620
likhet med unc-93 homolog B1 (C. elegans), transkripsjon variant 1 (LOC255620), mRNA
ILMN_1669393
GGT1
Gamma-glutamyltransferase en
ILMN_1685798
MAGEA6
melanom antigen familie A, 6
ILMN_3269395
GGT2
Gamma-glutamyltransferase 2
ILMN_1669833
SH2B2
SH2B adapter protein 2
ILMN_3238534
LOC100133817
Hypotetisk protein LOC100133817
ILMN_2099315
TRPM8
Transient reseptor potensielle kasjon kanal, underfamilien M, medlem 8
ILMN_3298065
LOC729195
Ligner apikale tidlig endosomal glykoprotein
ILMN_1717726
FLJ43752
Long intergeniske ikke-proteinkodende RNA 336
ILMN_1670452
ANKRD20A1
Ankyrin gjenta domene 20 familie, medlem A1
ILMN_3201060
LOC100132655 <.no> Hypotetisk protein LOC100132655
ILMN_3282829
LOC727913
Ligner iduronat 2-sulfatase (Hunter syndrom)
ILMN_2339691
SYVN1
Synovialt apoptose hemmer en , synoviolin
ILMN_1785549
SLC30A2
oppløst stoff carrier familie 30 (sink transporter), medlem 2
ILMN_3191898
LOC100129630
Hypotetisk LOC100129630
ILMN_1704204
LOC642204
Ankyrin gjenta domene som inneholder protein 26 lignende
ILMN_1682280
LOC647753
Hypotetisk protein LOC647753
ILMN_3201944
LOC646438
Hypotetisk LOC646438
ILMN_2233314
SPANXA1
Sperm protein forbundet med kjernen, X-bundet, familiemedlem A1
ILMN_3305980
NS3BP
NS3BP
ILMN_1747850
CRIM2
Kielin /chordin-lignende protein
ILMN_1700590
LOC645590
likhet med cAMP-avhengig protein kinase type i-beta regulatorisk subenhet
ILMN_1766316
FLJ32679
Golgin lignende hypotetisk protein LOC440321
ILMN_1890741
Hs
0,552561
bukspyttkjertelen holme cDNA-klon hbt09690 3, mRNA sekvens
ILMN_3308255
MIR33A
mikroRNA 33a
ILMN_1815716
LMLN
Leishmanolysin-lignende (metallopeptidase M8 familie)
ILMN_1654945
DNMT3A
DNA (cytosin-5 -) - metyltransferase 3 alpha
ILMN_2256050
SERPINA1
Serpin peptidase hemmer, klade A (alfa-1 antiproteinase, antitrypsin), medlem 1
ILMN_1759487
EGFLAM
EGF-aktig, fibronektin type III og laminin G domener
ILMN_1760410
LOC653086
likhet med RAN-bindende protein 2-aktig en isoform 2
ILMN_1668969
MIXL1
Bland sammen lignende homeobox
ILMN_3279757
LOC100132532
Hypotetisk protein LOC100132532
ILMN_1715372
CAMKK1
kalsium /calmodulin-avhengig protein kinase kinase 1, alfa
ILMN_1731370
Alle forfattere lese og godkjent den endelige manuskriptet.
 Oppsøk en gastroenterolog for kronisk diaré
Alle har en og annen anfall av diaré. Det kan være stress eller noe du spiste; Men hva om diaréen din ikke forsvinner? Hvis du opplever diaré konsekvent i flere uker, har du å gjøre med kronisk diaré.
Oppsøk en gastroenterolog for kronisk diaré
Alle har en og annen anfall av diaré. Det kan være stress eller noe du spiste; Men hva om diaréen din ikke forsvinner? Hvis du opplever diaré konsekvent i flere uker, har du å gjøre med kronisk diaré.
 De 4 beste tingene å vite om probiotika
Du vet sikkert om probiotika. De er de små, vennlige insektene som støtter en balansert og sunn tarm. Det du kanskje ikke vet er at fordelene deres går langt utover tarmhelsen!* Kroppen din er full a
De 4 beste tingene å vite om probiotika
Du vet sikkert om probiotika. De er de små, vennlige insektene som støtter en balansert og sunn tarm. Det du kanskje ikke vet er at fordelene deres går langt utover tarmhelsen!* Kroppen din er full a
 Mikrobiomtesting:Hva er dysbiose og sjekker du etter det?
Når tarmmikrobiomet er ubalansert, kan det skape problemer for fordøyelsen og den generelle helsen. Eksperter kaller det dysbiose, og her er det du trenger å vite om det. Hver uke, ledet av Tania, sv
Mikrobiomtesting:Hva er dysbiose og sjekker du etter det?
Når tarmmikrobiomet er ubalansert, kan det skape problemer for fordøyelsen og den generelle helsen. Eksperter kaller det dysbiose, og her er det du trenger å vite om det. Hver uke, ledet av Tania, sv