Abstract
Nedbryting av interstitiell celler av Cajal (ICC) er sertifisert i magen av diabetespasienter . Selv om elektroakupunktur (EA) på ST36 er en effektiv behandling for å regulere gastrisk motilitet, mekanismene for EA på ST36 på gastrisk tømming og nettverk av ICC gjenstår å bli belyst. Målet med denne studien var å undersøke effekten av EA på magetømming og på endringer i ICC-nettverk. Rotter ble randomisert til kontrollgruppen, diabetiske rotter (DM), diabetiske rotter med humbug EA (DM + SEA), diabetiske rotter med lav frekvens EA (DM + LEA) og diabetiske rotter med høy frekvens EA grupper (DM + HEA). Ekspresjonen av c-kit i hvert lag av mageveggen ble bestemt ved Western blotting. Spredning av ICC ble identifisert ved immunolabeling av c-kit og Ki67 som apoptose av ICC ble undersøkt ved TUNEL-farging. Resultatene var som følger: (1) Magetømming ble sterkt forsinket i DM-gruppen, men akselerert i LEA og HEA gruppe, spesielt i LEA-gruppen. (2) ekspresjon av c-kit i hvert lag ble redusert tilsynelatende i DM-gruppen, men også oppregulert i LEA og HEA gruppe. (3) Rikelig proliferert ICC (c-kit + /Ki67 +) dannelse av buskaktige nettverk med c-kit + celler ble observert i den LEA og HEA gruppe, mens de apoptotiske celler (c-kit + /TUNEL +) ble nesten ikke fanges opp i LEA og HEA gruppe . Kollektivt, lav og høy frekvens EA på ST36 redde skadede nettverk av ICC ved å hemme apoptose og styrke spredning i magen av diabetiske rotter, noe som resulterer i en forbedret gastrisk tømming
Citation. Chen Y, Xu JJ Liu S, Hou XH (2013) elektro på ST36 lindrer magetømming og redder Networks av interstitiell celler av Cajal i magen av diabetiske rotter. PLoS ONE 8 (12): e83904. doi: 10,1371 /journal.pone.0083904
Redaktør: Yvette Tache, University of California, Los Angeles, USA
mottatt: 09.08.2013; Godkjent: 08.11.2013; Publisert: 31.12.2013
Copyright: © 2013 Chen et al. Dette er en åpen-tilgang artikkelen distribueres under betingelsene i Creative Commons Attribution License, som tillater ubegrenset bruk, distribusjon og reproduksjon i ethvert medium, forutsatt den opprinnelige forfatteren og kilden krediteres
Finansiering:. Arbeidet ble delvis støttet med tilskudd fra Natural Science Foundation National of China (No. 30670775, nr 81270458). Ingen ekstra ekstern finansiering mottatt for denne studien. Finansiører hadde ingen rolle i studiedesign, datainnsamling og analyse, beslutning om å publisere, eller utarbeidelse av manuskriptet
Konkurrerende interesser:.. Forfatterne har erklært at ingen konkurrerende interesser eksisterer
Innledning
typer gastrointestinale motilitetsforstyrrelser spenner fra funksjonell dyspepsi til gastroparese alvorlig påvirke livskvaliteten til pasienter med tiår av diabetes mellitus [1]. Gastroparese, karakterisert ved forsinket gastrisk tømming, oppstår opp til 30% av individer med type 1 diabetes [2]. Selv om ulike terapeutiske intervensjoner inkludert legemiddelbehandling og noen ikke-medikamentelle behandlinger har vært prøvd, de helbredende virkningene for gastroparese fortsatt utilfredsstillende.
Akupunktur har blitt empirisk brukt til å behandle gastrointestinale forstyrrelser i orientalske land i tusenvis av år. Elektro (EA), som er en modifikasjon av tradisjonell akupunktur, er mer egnet for kliniske og forskningsgrunninnstillinger for dens reproduserbarhet. Zusanli plakater (ST36) er en vanlig acupoint ofte brukt til å behandle gastrointestinale sykdommer. I de senere årene, har effekten av EA på ST36 på gastrisk motilitet blitt undersøkt i en rekke studier. For dyr, EA i ST36 betydelig øket gastrisk motilitet i bevisste rotter og forbedret forsinket gastrisk tømming i STZ-indusert diabetiske rotter [3], [4]. Videre i mennesker, EA på ST36 akselerert ventrikkeltømming og lettet dyspeptiske symptomer på funksjonell dyspepsi pasienter og diabetiske individer med gastroparese [5], [6]. Selv i disse studiene kun lavfrekvente EA ble vedtatt, ble høyfrekvent EA også rapportert å fremme esophageal og distal kolon motilitet i dyr [7], [8]. Dermed blir effekten av forskjellig frekvens EA på gastrisk motilitet og de underliggende mekanismene trekke folks oppmerksomhet. Mellomliggende celler av Cajal (ICC), veletablerte som pacemakere som genererer og forplanter langsomme bølger, har fått betydning i regulering gastrointestinal motilitet over flere tiår [9]. Tre undergrupper av ICC inkludert intramuskulær ICC (ICC-IM), myentericus ICC (ICC-MIN) og submucosal ICC (ICC-SM) form intakte nettverk i mageveggen [10]. Nettverk av ICC er ikke statiske selv under fysiologiske betingelser; apoptose og transdifferensiering føre til ICC tap, mens ICC også kan gjenopprettes ved spredning, påfyll fra stamceller og øker overlevelse [11], [12]. Omfattende studier har dokumentert at tap og skader av ICC forårsaket uordnede gastrointestinal motilitet i dyremodeller og pasienter med diabetes [13] - [15]. Nylig, videre studier rapporterte at EA tilrettelagt økningen av colonic ICC i sakte transitt forstoppelse rotter og rotter etter enteroenterostomy [16], [17]. Vår fersk studie viste også at EA på ST36 kunne gi nedsatt ICC i tykktarmen av diabetiske rotter [18]. I mellomtiden er det opptatt av at EA fremmet proliferasjon og apoptose inhibert av hjerneceller i hippocampus [19], [20]. Men det er en blank om virkningene av EA på ST36 på ICC i diabetisk gastroparese og mekanismen om spredning og apoptose av ICC er involvert Derfor målene med denne studien var å:. 1) evaluere virkningene av forskjellig frekvens EA på gastrisk tømming; 2) undersøke effektene av EA på ST36 på ICC nettverk i diabetiske rotter; 3) sertifisere om apoptose og spredning av ICC var involvert. Etikk uttalelse Rottene fikk menneskelig omsorg og denne studien ble utført strengt i samsvar med anbefalingene i guide for omsorg og bruk av forsøksdyr av National Institutes of Health, med alle operasjoner som er godkjent av de etiske retningslinjene fra Animal Care og bruk komité Tongji medisinske høyskole, Huazhong University of Science and Technology. Dyr Sprague-Dawley rotter veide 250-300 g ble valgt i denne studien. Rotter ble hentet fra Experiment Animal Center of Tongji Medical College (Huazhong University of Science and Technology, Wuhan, Kina) og holdt under normale laboratorieforhold (22 ° C, 12/12 timers lys-mørke-syklus) med fri tilgang til mat og sterilt vann. Etter en uke med adaptiv fôring, ble dyrene offisielt vedtatt i våre eksperimenter. Etter faste over natten, men fri tilgang til vann, ble modellene etablert med et enkelt intraperitoneal injeksjon av streptozotocin i rotter (STZ, 60 mg /kg, dvs. 1 ml /100 g kroppsvekt, Alexis biokjemikalier, San Diego, CA, USA), fersk oppløst i 20 mM citratbuffer løsning (pH 4,5, Sigma, St. Louis, MO, USA) . Kontrollgruppen ble injisert med et likt volum av sitratbuffer. Alle rottene ble holdt i den samme tilstand, med vannmangel i 4 timer, men ad libitum Rotter ble randomisert inn i fem grupper (10 rotter hver gruppe), inkludert kontroll , diabetiker gruppe (DM), diabetiske rotter med humbug EA gruppe (SEA, bare akupunktur uten elektrisk strøm, 30 min /d), diabetiske rotter med lav frekvens EA gruppe (LEA, 10 Hz, 1-3 mA, 30 min /d ) og diabetiske rotter med høy frekvens EA gruppe (HEA, 100 Hz, 1-3 mA, 30 min /d). EA ble gjennomført på acupoint ST36 med en elektrisk stimulator (G6805-2A; Shanghai Lam Medical Instrument Factory, Shanghai, Kina) hverdagen i 8 uker. Den elektriske strømmen ble etablert i henhold til litt skjelvende i underekstremitetene. For å oppnå det formål å eliminere innflytelsen av stress, ble dyrene lov til å bevege seg fritt i sitt eget foringsrør under EA prosedyren. Etter suksessiv intervensjon i 8 uker, ble rottene i hver gruppe testet med hensyn til gastrisk tømming. Deretter ble de ofret, og eksemplarer av antrum ble nøye oppnådd. Hver prøve ble delt i flere stykker, er en forholdsvis liten en lagret ved -80 ° C for Western blot-analyse etter lag ferskt frittliggende og et stykke ble anbragt i 2,5% glutaraldehyd for elektronmikroskopi, og resten ble løst i Zamboni s fiksativ (2- % paraformaldehyd og 1,5% mettet picrinsyre løsning i 0,1 M fosfatbufferoppløsning (PBS, pH 7,4) for immunofluorescens farging. Måling av gastrisk tømming mage~~POS=TRUNC tømming~~POS=HEADCOMP ble utført ved en modifikasjon som beskrives tidligere [21]. test~~POS=TRUNC måltid inneholdende fenol-rødt (0,5 mg /ml) som indikator og karboksymetylcellulose (15 mg /ml) ble stadig omrørt og deretter holdt ved 37 ° C. Etter at mat-belastede over natten, ble rottene mottok 2 ml fenol rød måltid administreres via en sonde nål. Tretti minutter senere ble dyrene hurtig avlivet ved cervikal dislokasjon. hele magen ble fjernet forsiktig etter ligering ved cardia og pyloriske, og deretter magen var åpen og dens innhold ble helt over i et reagensglass og vasket med 40 ml destillert vann. Ved slutten av eksperimentet, ble NaOH-oppløsning (20 ml, 1 M) tilsatt til hvert rør for å utvikle den maksimale intensiteten av fargen. Absorbansen til prøven avlest ved 560 nm med et spektrofotometer (Hitachi U-2900) var å gjenspeile mengden av fenolrødt igjen i magen. Frekvensen av gastrisk tømming ble beregnet i henhold til følgende formel: Gastrisk tømming (%) = 100 x (1 - X /Y), X representerer absorbans av fenolrødt oppsamlet fra magen hos dyr avlivet 30 minutter etter testmåltidet. Y betegner absorbans av fenolrødt utvinnes fra magen hos kontrolldyr drept umiddelbart etter administrasjon av testmåltidet. Friske prøver av gastrisk antrum ble festet til en Sylgard tallerken og hvert lag ble frittliggende omhyggelig med skarp disseksjon etter fjerning av slimhinnen. Med hjelp av fine-spiss tang og mikro saks under et dissekere mikroskop, ble lag av ICC-SM, ICC-IM og ICC-MY oppnådd. Alle manipulasjoner ble utført på is for å unngå at proteintap. Og deretter prøver av ICC-SM, ICC-IM og ICC-MIN ble lagret ved -80 ° C. Frisk-frosne prøvene ble knust i cellesuspensjonen og homogenisert i RIPA-buffer (Upstate, Temecula, CA , USA) med proteasehemmer (Sigma Chemical Co., St. Louis, MO, USA). Blandingen ble sentrifugert ved 12000 g i 10 min ved 4 ° C, og supernatantene ble samlet opp som det totale protein. Proteinkonsentrasjonen ble bestemt ved Bradford-metoden. Deretter ekvivalenter av 50 mg av ekstraherte proteiner ble gradvis separert ved 10% natriumdodecylsulfat polyakrylamid gelelektroforese (SDS-PAGE) og overført til PVDF-membran (Millipore, Bedford, MA, USA). Fulgt blokkere ikke-spesifikke bindingssteder med 5% fettfri tørrmelk i Tris-bufret saltvann inneholdende 0,1% Tween 20 (TBST) ved romtemperatur i 1 time, ble disse membranene inkubert med primært antistoff til anti-c-kit (1:200, Santa Cruz Biotechnology, Inc.) over natten ved 4 ° C sammen med kanin-anti-mus Actin (1:400, Santa Cruz Biotechnology, Inc.) tjente som indre kontroll. Etter tre vaskinger med TBST, membranene inkubert med HRP-koblet sekundært antistoff (HRP-bundet kanin-anti-geit og HRP-bundet geit-anti-mus, 1:5000) i 1 time ved romtemperatur. Fulgt med tre vaskinger med TBST, ble båndene visualisert ved kjemisk reaksjon med forbedret kjemiluminescens middel (Amersham Pharmacia Biotech, Piscataway, NJ) og den blot ble utsatt for autoradiografi. Densitometry analyse ble utført av Antall Én programvare (Bio-Rad tekniske avdeling, versjon 4.6.2). For hele monterte flekker forberedelser, magen ble åpnet og renset flere ganger med PBS. Gastric antrum ble festet til en Sylgard rett og strukket til 150% av sin opprinnelige størrelse, og deretter fast med Zamboni er fiksativ i 24 timer ved romtemperatur. De fikserte vev ble vasket med PBS og deretter mukosa ble fjernet ved skarp disseksjon. ICC-SM finne på submucosal overflaten av den sirkulære muskellaget, ICC-IM intermingling blant glatte muskelceller og ICC-MY sammen med myentericus nerve plexus ble nøye utarbeidet etter fine spisse pinsett og mikro saks under et dissekere mikroskop. De farging rutiner for å avdekke ICC nettverk og spredning har tidligere blitt beskrevet [22]. Ki67, et kjerneprotein uttrykt i løpet av alle de aktive faser av celle og forsvant i den hvilende celle, har blitt anvendt for å påvise den proliferative ICC [23]. Etter vasking seks ganger med PBS i 1 time, ble vevene inkubert med 5% normalt bovint serum i PBS inneholdende 0,3% Triton X-100 (PBST) i 1 time ved romtemperatur og deretter inkubert med primært antistoff c-kit (1:100; Santa Cruz, CA, USA) og Ki67 (1:100; Abcam, Cambridge, UK) over natten ved 4 ° C. Etter skylling i PBST i 30 minutter, ble immunreaktiviteten påvist ved anvendelse av Alexa Fluor 594-konjugert sekundært antistoff (esel anti-geit IgG, 1:100) og Alexa Fluor 488-konjugert sekundært antistoff (esel anti-kanin IgG, 1:100) ved 37 ° C i 1 time, etterfulgt av skylling i PBS tre ganger i 30 min, og inkubert med DAPI (0,1 mg /ml; Sigma, St. Louis, MO, USA) i 30 minutter som en kjernefysisk teller flekk. Skylt i PBS to ganger, vev var utfoldet på et lysbilde. En konfokal laser scanning mikroskop (Nikon, Japan) ble brukt for å undersøke de immunolabeled vev. For å inkludere alle nivåer av positivt fargede celler og prosesser, ble den optimale Z stabling av confocal bilder satt til 5 mikrometer intervaller. apoptose av ICC ble oppdaget med TUNEL merking, en apoptose deteksjon Kit (Roche, Tyskland). Hel-mount forberedelser ble inkubert med primært antistoff c-kit (1:100, Santa Cruz, CA, USA) over natten ved 4 ° C og sekundært antistoff (Alexa Fluor 594 esel anti-geit IgG, 1:100) ved 37 ° C i 30 minutter. Under en fuktet og mørke forhold, ble vevene inkubert med TUNEL reaksjonsbuffer (merking løsning: enzymløsning = 09:01) ved 37 ° C i 1 time. Etter å ha vasket tre ganger i 30 minutter for å fjerne ukoblet fluorescein-dUTP og inkubert med DAPI som en kjernefysisk teller flekk, ble prøvene observert under en konfokal laser scanning mikroskop. Når mucosa fjernet, ble prøver av antrum neddykket i et fiksativ oppløsning av 2,5% glutaraldehyd ved 4 ° C i 2 timer. Deretter ble de renset med 0,1 M PBS to ganger i 20 min, post-fiksert i 1% OsO 4 (pH 7,4) i 1 time, dehydrert med gradert alkohol, klaret i propylenoksyd og innleiret i Epon ved hjelp av flate støpeformer. Ultratynne seksjoner ble oppnådd med en ultramicrotome (Leica ULTRACUTU, Tyskland), og farget med bly citrate i 10 minutter før visning med en transmisjonselektronmikroskop (Tecnai G 212, FEI, Nederland). for immunfluorescens farging analyse, mage antrum fra hvert forsøksdyr ble samplet på samme sted i magen. ICC ble gjenkjent av c-kit immunreaktivitet og tilstedeværelsen av tallrike cellulære prosesser. Prolifererende og apoptotisk ICC ble identifisert ved c-kit immunoreaktivitets og Ki67 /TUNEL farget kjerne omslutter med c-kit positiv membranstrukturer. Ti tilfeldige felt (× 200 forstørrelse, 0,2607 mm 2) per hel-mount forberedelse ble tatt for fotografier av c-kit positive celler, c-kit + /Ki67 + og c-kit + /tunel + merkede celler. Disse fotografiene ble også anvendt for å vurdere tykkelsen av lagene i hver stabel ved hjelp av DAPI merkede kjerner som en markør av de øvre og nedre grenser av lagene. Tallene for c-kit positive celler, ble c-kit /Ki67 og c-kit /tunel dobbelt merkede celler telles med bilde-ProPlus 5.0 (Media Cybernetics). Alle data ble uttrykt som gjennomsnitt ± SEM. Minst ti visjoner ble tatt i hvert dyr. Analysen av variansen (ANOVA) og t-test ble brukt for å sammenligne forskjellene mellom ulike grupper. P-verdi mindre enn 0,05 ble tatt som statistisk signifikant forskjell. Kroppsvekt og blodsukkernivået Etter 8 uker, skjedde dødsulykker i DM-gruppen (to rotter) og i SEA gruppen (en rotte). Rottene ble screenet for den samme utgangsvekten mellom gruppene (Tabell 1). Ved slutten av en, 4 og 8 uker, ble kroppsvekten av DM gruppen signifikant redusert sammenlignet med kontrollen ( P Som vist i tabell 2, ble heller ikke vist forskjeller i referanseblodsukkernivået mellom gruppene. De diabetiske modeller ble etablert suksess med høyt blodsukker i DM-gruppen sammenlignet med kontrollgruppen på slutten av en, 4 og 8 uker (alle P Fig.1 viser effekten av EA i ST36 på gastrisk tømming i forskjellige grupper. Den gastriske tømming av DM-gruppen ble betydelig forsinket i motsetning til kontrollgruppen ( P fra Fig.2, uttrykket av c-kit i hvert lag av mageveggen ble vurdert. I ICC-IM, c-kit i DM-gruppen ble åpenbart redusert sammenlignet med kontrollgruppen (0,47 ± 0,04 vs 0,69 ± 0,05, P Effekter av EA på ICC nettverk vekslinger av ICC nettverk spesielt ICC morfologi og tetthet ble vist i figur 3. Det var ingen signifikant forskjell av lag tykkelse mellom gruppene. Forbundet med glatte muskelceller, c-kit immunoreaktivitet for ICC-IM ble observert i muskellagene, som viser at ICC-IM hadde to polare prosesser som danner et celledelt nett i kontrollgruppen. I DM gruppen ble de lange prosesser av ICC-IM nedsatt tilsynelatende og tettheten av c-kit positiv celle ble redusert til ca. 50% av normal kontroll ( P ICC-MY svingete rundt myentericus plexus lå mellom langsgående og sirkulære glatte muskellagene. Multipolar ICC-MY med mange grener som danner en intakt mobiltelefonnettet var tydelig viste i kontrollgruppen. I DM-gruppen, c-kit + celler dukket opp med slanke celle organer og oppsprukne prosesser, og tettheten av ICC-MY var åpenbart redusert ( P apoptose av ICC ble oppdaget av TUNEL merking for å avgjøre om apoptose var involvert i mangel av ICC i diabetes (Fig.4). Av hele montering preparater for kontroll, ble noen få kjerner av ICC farget ved TUNEL-metoden i muskellaget, myenterisk lag og submucosa lag. Rikelig med c-kit + /TUNEL + ICC-IM, ICC-MY og ICC-SM ble observert i DM-gruppen (18,48 ± 1,88 mm -2, 26.51 ± 1.87 mm -2 og 12.03 ± 1.27 mm -2, henholdsvis). C-kit + mobilnettverk var ufullstendige og apoptotisk ICC presentert som forkortet prosesser med færre avdelinger. Store mengder c-kit + /tunel + celler ble også flom i alle lag av SEA gruppen. Under gjennomføringen av LEA og HEA, ble apoptotiske kit + celler neppe funnet i ICC-IM (1,79 ± 0,33 mm -2 og 2,05 ± 0,30 mm -2), ICC-MY (2,24 ± 0,40 mm -2 og 2,48 ± 0,40 mm -2) og ICC-SM (1,09 ± 0,25 mm -2 og 1,39 ± 0,20 mm -2). Mobilnettverk avslørt av c-kit farging viste høy tetthet og tykkelse av cellulære prosesser i LEA og HEA gruppe. Med tanke på økt antall ICC i LEA og HEA gruppe, Ki67-positive ICC ble påvist å avsløre spredning av ICC i de tre lagene (fig.5). I kontrollgruppen, et antall av c-kit /Ki67 dobbeltmerkede celler ble observert i ICC-IM (8,82 ± 1,02 mm -2), ICC-MY (10,64 ± 0,92 mm -2) og ICC -SM (4.67 ± 0.96 mm -2). Antallet ICC med brukket grenene var bemerkelsesverdig redusert og nettverkene var svært sparsommelig med få prolifererende celler både i DM og SEA gruppen. Imidlertid ble i LEA og HEA gruppe, c-kit /Ki67 dobbelt merkede celler i ICC-IM preget av tett tilstøtende celle organer og relativt bipolar prosesser, og den gjennomsnittlige tettheten nådd til 18.37 ± 1.60 mm -2 og 15,27 ± 1,81 mm 2 (begge P Effekter av EA på ultra endringer av ICC Typiske ultra kjennetegn ved ICC ble avslørt av transmisjonselektronmikroskopi (figur 6). I kontrollgruppen, ICC-IM og ICC-Mitt rike av velutviklede celleorgan utvidet lange prosesser som inneholder mitokondrier, og grov og glatt endoplasmatisk retikulum. Polymorfe ICC-SM plassert på submucosal overflate av den sirkelrunde muskellaget viste høyere elektron tett cytoplasma, tallrike mitokondrier og endoplasmatisk retikulum. I DM gruppen, ble en dramatisk reduksjon i antallet ICC med skadede ultrastrukturelle egenskaper undersøkt ved elektronmikroskopi. Disse cellene med hovne mitokondriene, forstyrret cristaed og kondensert kromatin viste ufullstendig membran og forkorte prosesser. Disse endringene ble også sett i SEA gruppen. I LEA og HEA gruppe, antall ICC var betydelig økt og de koblet tett med nervefibre og glatte muskelceller som danner synaptiske struktur og gap veikryss. Clear mitokondrier crest, ryddig distribuert grove endoplasmatiske retikulum og ribosomale partikler ble ansett for å være klassiske trekk ved ICC-IM, IM-MY og ICC-SM. I denne studien har vi funnet at både lav og høy frekvens EA ved ST36 undertrykket apoptose av ICC og aktivert spredning av ICC i magen hos STZ-indusert diabetiske rotter. Som et resultat ble redning av ICC-nettverk forbundet med en hindrende virkning på forsinket gastrisk tømming. som en effektiv fremgangsmåte med færre bivirkninger, EA kombinert med akupunktur og elektrisk strøm stimulering i stedet for manuelle manipulasjoner blir gradvis akseptert av Vestlige land. ST36 er en av de hyppigst brukte poeng til å behandle gastrointestinale forstyrrelser, med indikasjoner inkludert: smerter i epigastriet, kvalme, oppkast, dårlig appetitt og abdominal oppblåsthet. Samtidig har vår tidligere studie indikerte EA på ST36 kan øke colonic propulsive bevegelse, men ikke observert i gruppen med EA på ikke-akupunkturpunkter. Forsinket ventrikkeltømming er en felles karakter av diabetespasienter med gastroparese og også en av de største faktorene som bidrar til symptomer på gastroparese. I løpet av de siste tiårene har eksperimenter i bevisste rotter viste at lavfrekvens EA (10 Hz) betydelig akselerert gastrisk tømming [3], [24], [25]. I tillegg, 100 Hz EA var effektiv til å forårsake frigjøring av dinorphin og serotonin for å indusere analgesi [26]. Selv om høye frekvens EA (100 Hz) ble også rapportert å fremme esophageal og distal colon motilitet hos dyr [7], [8], virkningene av 100 Hz EA på gastrisk motilitet knapt har undersøkt. Dermed ble parametere (10 og 100 Hz) fra EA valgt ifølge foreløpige eksperimenter som indikerte stimulerende effekt av gastrointestinal motilitet. Våre resultater viste at både lav frekvens EA (10 Hz) og høyfrekvens-EA (100 Hz) ved ST36 betydelig forbedret forsinket gastrisk tømming i diabetiske rotter, og resultatet viser også at virkningen av lav frekvens EA var mer dramatisk enn den for høy frekvens EA. Tallrike studier har vist at ICC spiller en viktig rolle i reguleringen av mage peristaltikk, en viktig determinant for gastrisk tømming [27], [28]. Som ICC finnes i ulike lag av gastrisk antrum i form av nettverk in vivo det er bemerkelsesverdig at balansen av ICC-nettverk er i hovedsak bestemt av apoptose og proliferasjon av ICC [11], [12]. Derfor ble det mulig mekanisme på gastrisk motilitet og effektene på ICC nettverk av EA på ST36 utforsket gjennom å detektere endringer av spredning og apoptose av ICC. Nylig er apoptotisk celledød rapportert å være en pågående prosess med ICC nettverk og graden av apoptose i sunn kolon indikerer at disse cellene må være kontinuerlig regenereres å opprettholde intakte nettverk [12]. Dessuten resultater fra voksne marsvin indikerte at intestinal ischemi og reperfusjonsskade førte til en reduksjon i antall av ICC ved apoptose som kan bidra til gastrointestinale motilitetsforstyrrelser [30]. Videre er en fersk rapport viste at EA terapi reduserte apoptotiske prosentandel av substantia nigra celler i Parkinson rotter [31]. Denne studien har klart formulert at apoptotisk ICC oppdaget av tunel metodene var flom i ICC nettverk av diabetisk rotte magen og spekulert i at uttømming av ICC-nettverk kan også føre til mage motilitetslidelser. Men gjennom intervensjon med lav og høy frekvens EA stimulering, TUNEL + kjernen var nesten ikke til å finne, noe som tyder på apoptose av ICC ble redusert til et lavt nivå som det normale. spredning er også en annen hovedregulator for å opprettholde nettverk av ICC. Tidligere resultater har vist at proliferasjonen var involvert i utvinning av ICC når det ble tapt i voksne dyr [32], [33].
Materialer og metoder
Diabetic modell
foring. En uke etter injeksjonen, ble diabetes bekreftet i forskjellig tid ved å måle glukose i en dråpe blod erholdt ved punktering av haletipp fartøyet. En blodglukosenivå ≥16.7 mmol /l ble tatt som en standard for vellykket modell. Blodsukkeret ble siktet punktvis før injeksjon og første, fjerde og åttende uke etter injeksjonen. Videre, vekttap, polydipsi og polyuri var flere indikatorer for å bevis den vellykkede tilskyndelse av diabetisk rotte.
Eksperimentelle protokoller
Western blotting
immunfluorescens studier
Elektronmikroskopi
Statistisk analyse
Resultater
= 0,009, 0,000 og 0,000). Det var ingen signifikant forskjell mellom SEA og DM-gruppen ( P
> 0,05). Men i motsetning til DM gruppen, ble kroppsvekten forhøyet i LEA gruppen ved 8 uker ( P
< 0,001) og HEA gruppe på 4 og 8 uker ( P =
0.003, P
. < 0,001 separat)
< 0,001). I mellomtiden, blodsukkernivået i havet, Lea og HEA gruppene ble ikke endret seg vesentlig i forhold til DM-gruppen (alle P
> 0,05).
Effekter av EA på ventrikkeltømming
= 0,012). Sammenlignet med DM gruppe, SEA ikke hadde noen signifikant effekt på magetømmingen ( P
= 0,338), men LEA og HEA markert akselerert forsinket tømming fra 43,96 ± 5,02% til 69,72 ± 3,02% og til 59.06 ± 3,70% ( P
< 0,001 og P
= 0,013, henholdsvis). Sammenligning mellom LEA og HEA gruppe, ble statistisk signifikans oppnådd ( P
= 0,04), som representerer at effekten av LEA åpenbart overskredet som HEA.
Effekter av EA på uttrykket av c -kit
= 0,001). Motsatt LEA og HEA dramatisk forbedret uttrykk for c-kit sammenlignet med DM-gruppen (0,65 ± 0,04 vs 0,47 ± 0,04, P
= 0,004; 0,66 ± 0,03 vs 0,47 ± 0,04, P
= 0,002, henholdsvis). I ICC-MY, ble ekspresjonen av c-kit betydelig redusert i DM gruppe i motsetning til kontrollgruppen (0,57 ± 0,04 vs 0,83 ± 0,05, P
< 0,001). Men signifikant forskjell ble begge oppnådd i LEA og HEA gruppen sammenlignet med gruppen DM (0,78 ± 0,05 vs 0,57 ± 0,04, P
= 0,002; 0,76 ± 0,04 &Co. 0,57 ± 0,04, P
= 0,004, henholdsvis). I ICC-SM, ble c-kit av DM gruppen tydeligvis minsket på basis av normalt nivå (0,56 ± 0,03 vs 0,77 ± 0,04, P
= 0,001). Konsekvent, Lea og HEA åpenbart økt uttrykk av c-kit sammenlignet med DM-gruppen ( P
= 0,003 og P
= 0,007). Men ingen signifikant forskjell ble funnet mellom SEA og DM-gruppen i ICC-IM, ICC-MY og ICC-SM ( P
= 0,597, P
= 0,853 og P
= 0,172).
< 0,001). Sammenlignet med DM gruppe, ble ingen signifikant endring bemerket i SEA-gruppen. Men en nesten intakt celledelt nett ble visualisert i LEA og HEA gruppe og tettheten av c-kit positiv ICC-IM nesten gjenopprettet til det normale nivå (både P
< 0,001, sammenlignet med gruppen DM ).
< 0,001). I SEA gruppe, tettheten av ICC-MIN ble ikke signifikant endret i forhold til DM gruppe ( P
= 0,762). Men en nesten intakt mobilnettverk eksisterte rundt myentericus plexus og ICC-MY tetthet delvis gjenfunnet i LEA og HEA gruppe (begge P
< 0,001, sammenlignet med DM-gruppen)
<. h3> Effekter av EA på apoptose av ICC
Effekter av EA om spredning av ICC
< 0,001). Tilsvarende rikelig c-kit + /Ki67 + ICC-MY i LEA og HEA gruppe (begge P
< 0,001) danner intakte nettverk med c-kit + celler utført en runde cellekroppen og lange slanke prosesser. I ICC-SM, ledsaget av et intakt nettverk av ICC, tettheten av prolifererende celler i LEA og HEA gruppe var forholdsvis høyere i motsetning til DM gruppen ( P
= 0,016 og P
= 0,003).
Diskusjoner
, endringer av ICC nettverk er valuably å bli vurdert. Reduksjon i antall og morfologiske endringer av ICC-MY og ICC-IM i magen av diabetiske mus ble vist i en tidligere studie [14]. Tilsvarende ble en markant tap av ICC-IM og ICC-SM i antrum av STZ-induserte diabetiske rotter observert, men ICC-MY ble ikke signifikant påvirket med unntak av et tap av forbindelser med nerve strukturer [15]. Disse forskjellene av funnene i disse to studiene ble sannsynligvis knyttet til dyrearter. Fra en studie av 28 pasienter med gastroparese, ble et redusert antall ICC funnet i plexus myentericus med full tykkelse antrumbiopsier fore [29]. Våre resultater viste at ICC-IM, ICC-MY og ICC-SM i antrum av STZ-induserte diabetiske rotter ble dramatisk redusert basert på teknikken av western blotting og immunfluorescens farging, og fremme skadet strukturen av ICC ble vist av transmisjonselektronmikroskopi . Heldigvis er det rapportert at lavfrekvent EA forbedrer uttrykk for ICC i tykktarmen av langsomme transitt forstoppelse rotter [17]. I tillegg har vår tidligere studie antydet at både lav og høy frekvens EA gjenoppretter uttrykk for ICC i kolon av diabetiske rotter [18]. Men i disse studiene, de undersøker endring av ICC som en helhet, og virkningene av forskjellig frekvens EA på ICC i magen er ikke blitt klarlagt. Dermed blir effekten av forskjellig frekvens EA på tre undertyper av ICC i magen inkludert ICC-IM, ICC-MY og ICC-SM var fokus for denne studien. Våre resultater viste at uttrykket av ICC i de tre lagene ble tydelig reddet av lav og høy frekvens EA, noe som tyder på at lav og høy frekvens EA renovert nettverk av ICC-IM, ICC-MY og ICC-SM og videre kan bidra til en gjenvunnet gastrisk tømming.
 Hva er det beste kostholdet for noen med ulcerøs kolitt?
Hva er ulcerøs kolitt? Ulcerøs kolitt er en kronisk tilstand der tykktarmen og endetarmen ofte er betent. Den beste dietten for noen med ulcerøs kolitt er en som inkluderer magert protein, lav-fibe
Hva er det beste kostholdet for noen med ulcerøs kolitt?
Hva er ulcerøs kolitt? Ulcerøs kolitt er en kronisk tilstand der tykktarmen og endetarmen ofte er betent. Den beste dietten for noen med ulcerøs kolitt er en som inkluderer magert protein, lav-fibe
 Cystinuri – nyrestein
Steinene sammenlignet her i størrelse med en fjerdedel (en mynt på USD 0,25) kom fra nyrene til en ung kvinne med cystinuri, den vanligste defekten som er kjent ved transport av en aminosyre (en bygge
Cystinuri – nyrestein
Steinene sammenlignet her i størrelse med en fjerdedel (en mynt på USD 0,25) kom fra nyrene til en ung kvinne med cystinuri, den vanligste defekten som er kjent ved transport av en aminosyre (en bygge
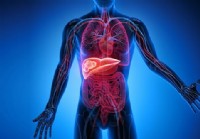 Leverkreft (hepatocellulært karsinom)
Fakta om leverkreft Leverkreft er ofte et resultat av kronisk leversykdom. De fleste som får leverkreft (leverkreft) får det i sammenheng med kronisk leversykdom. Forekomsten av hepatocellulær kreft
Leverkreft (hepatocellulært karsinom)
Fakta om leverkreft Leverkreft er ofte et resultat av kronisk leversykdom. De fleste som får leverkreft (leverkreft) får det i sammenheng med kronisk leversykdom. Forekomsten av hepatocellulær kreft