etäpesäkekasvainten kehittyminen ja organoidikappaleen mallinnus sotkea TGFBR2
syövän kuljettaja hajanainen mahasyövässä
tiivistelmä
tausta
Mahalaukun syöpä on toiseksi -leading syy maailmanlaajuiseen syöpäkuolemista, etäpesäkkeitä edustaa ensisijainen kuolinsyy. Kartoittaakseen kuljettajat osallistuvat onkogeneesiin ja kasvaimen kehittymisen, me tehtävä laaja Genomikartoituksen analyysi metastasointiin on hajanainen mahasyövän. Tämä edellyttää vertailua ensisijainen kasvain perinnöllinen hajanainen mahasyövän oireyhtymä proband ja sen toistumisen kuin munasarjojen etäpesäkkeitä.
Tulokset
Sekä primaarikasvaimen ja munasarjojen etäpesäkkeitä on yhteinen bialleelisten menettämisestä toiminta sekä CDH1
ja TP53
tuumorisuppressoreilla, osoittaa yhteinen geneettinen alkuperä. Vaikka primaarikasvaimen esittelee monistus fibroblastien kasvutekijän reseptori 2 (FGFR2
) geenin, etäpesäke erityisesti puuttuu FGFR2
vahvistusta, vaan on ainutlaatuiset bialleelisen korjauksilla transformoivan kasvutekijä-beeta-reseptori 2 (TGFBR2
) , mikä osoittaa erilaiset in vivo
kehittyminen on TGFBR2
-mutant metastaattisen klonaalisen populaation tässä potilasryhmässä. Kuten TGFBR2
mutaatioita ei ole aiemmin toiminnallisesti validoitu mahasyövän, me mallinnettu metastaattista potentiaalia TGFBR2 menetys hiiren kolmiulotteinen ensisijainen mahalaukun organoidikappaleen kulttuuriin. Tgfbr2
shRNA Knockdown sisällä Cdh1
- /-
; Tp53
- /-
organoids generoi invaasion in vitro
ja vankka metastaattisen tuumorigeenisyystesti in vivo
, vahvistaa Tgfbr2
etäpesäke suppressoriaktiivisuutta.
Johtopäätökset
Me dokumentoida metastasoitunut erilaistumista ja geneettinen heterogeenisyys hajanainen mahasyövän ja paljastaa mahdolliset metastaattisen roolia TGFBR2
menettämisestä toiminnon. Tueksi Tässä tutkimuksessa käytämme hiiren ensisijainen organoidikappaleen kulttuuri menetelmä pystyy pääpiirteittäin in vivo
metastaattisen mahasyövän. Kaiken kuvaamme yhtenäinen lähestymistapa tunnistaa ja toiminnallisesti vahvistaa otaksuttu syövän kuljettajien mukana etäpesäkkeitä.
Background
Worldwide, mahalaukun adenokarsinooma on neljänneksi yleisin maligniteetti ja toiseksi yleisin syy syöpäkuolemista miehillä ja naisilla. Perustuu erottuva histopatologinen piirteitä, mahalaukun adenokarsinooman on luokiteltu hajanainen ja suoliston alatyyppejä [1]. Mitä histopatologia, hajanainen syöpien ovat yleensä erilaistumaton, usein sinettisormus solu rengas ominaisuuksia ja invasiivisesti suodattuvat normaalin vatsan kudosta. Sitä vastoin suolen alatyypin on epiteelin ominaisuuksia ja muodostaa erillisiä kasvainmassojen samanlainen paksusuolen syöpä. Diffuusi mahasyöpä on korkeampi esiintyvyys etäpesäkkeitä ja yleisesti huonompi ennuste verrattuna suoliston alatyypin [2], [3]. Tällä hetkellä genominen analyysit hajakuormituksen mahasyövän ovat mukana pieni määrä näytteitä kuten tuore tutkimus Cancer Genome Atlas Project (TCGA) ja koko Genomikartoituksen tutkimus joukko hajanainen mahalaukun kasvaimet [4]. On kuitenkin olemassa vähän, jos lainkaan, tutkii että yksityiskohtaisesti metastaattinen kehittymistä mahasyöpä; etäpesäkkeitä ovat tyypillisesti poissa suuren mittakaavan genominen syövän tutkimuksista, kuten TCGA. Kaiken kaikkiaan tiedetään vähän onkogeenisel- prosessia ja kasvainten kehittymistä metastasoituneen mahalaukun syövän huolimatta ensiarvoisen kliinistä merkitystä [5].
Perinnöllisen hajanainen mahasyövän (HDGC), ituradan mutaatioita CDH1
(eli E- kadheriinin) koituu 70% elinikäinen riski sairastua diffuusi mahasyövän [6], [7]. CDH1
tuumorisuppressorigeenin koodaa E-kadheriinin, läpäisevä glykoproteiini, joka välittää kalsiumista riippuvan solu-solu-adheesion. Muutoksia CDH1 toiminto vaikuttaa epiteelin-mesenkymaalitransitioon (EMT), joka on ollut mukana niin pelissä rooli kasvainten synnyssä. Tutkimukset sairastuneiden HDGC yksilöiden kasvaimia tarjoavat ainutlaatuisen mahdollisuuden määrittää olennainen kuljettajia hajakuormituksen mahasyövän yhteydessä CDH1
toimintakyvyn menetystä. Tukevat todisteet roolin CDH1
satunnaisissa hajanainen mahasyövistä voidaan mainita, että 50% sisältävät CDH1
mutaatioita tai hypermetylaatiota CDH1
promoottori [8], [9]. Tuore kokonainen Genomikartoituksen kyselyn hajanainen mahasyövän myös tunnistaa usein CDH1
mutaatioita yleisin kuljettaja tapahtuma [4]. TCGA mahasyöpä tiedot osoittavat myös tiheä somaattisten CDH1
mutaatioista [10]. Huomattavasti vähemmän tiedetään identiteettiä ja roolia yhdessä esiintyvät kuljettajat, jotka edistävät hajanainen mahalaukun etäpesäkkeitä.
Tässä me raportoimme tutkimuksen metastaattisen evoluutioprosessin heikossa mahasyövässä. Tavoitteena oli tunnistaa tunnettujen ja ehdokas ohjaimet, joissa kartoitetaan kasvainprogression aikana etäpesäkkeiden. Suoritimme laajan Genomikartoituksen analyysi ensisijainen mahakasvaimen ja etäpesäkkeiden yksilöstä, jolla on ituradan CDH1
mutaatio (kuvio 1), joka esitetään mahalaukun ensisijainen, sen jälkeen kun 3 vuotta, jonka etäpesäke vasemmalla munasarja. Ottaen huomioon nykyiset ituradan mutaatio CDH1
syöpä genomin vaatii vain toinen alleelista osuma kautta somaattisen geneettinen poikkeavuus, kuten ilmenee kasvaimen kyseisen yksilön. Koska alkuperäisen syöpä kuljettaja tapahtuma tunnetaan, Mendelin syöpä genomit tarjota harvinaisen ja erittäin informatiivinen `koe luonnon", joka antaa mahdollisuuden rajata somaattisten genetiikkaa etäpesäke. Genomikartoituksen analyysi sekä kasvainten paljasti näyttöä yhteinen alkuperä perustuu yhteisiin mutaatioita mutta suurempi genomista monimuotoisuutta nähdä sekä tasolla mutaatiot sekä laajaa alleeliset epätasapaino ja kopioi numero aberraatioita varten metatasis. Kuvio 1 Perhe ja kliinisen historian Mendelin hajanainen mahasyövän. Sukutaulu indeksin potilaan 525 (III-1) on kuvattu. Tuumorin tyyppi on mainittu värin mukaan lukien vihreä haimasyöpä, punainen hajanainen mahasyövän, ja keltainen rintasyövän. Potilaalla oli hänen ensisijainen mahasyövän iässä 37 vuotta. Kolme vuotta myöhemmin hän esitti kanssa vatsavaivoja. Varjoainetehostettu CT lantion tunnistettu vasen munasarja massa (keltainen ympyrä), joka vahvistettiin koepala olla hajanainen mahasyövän etäpesäkkeiden (eli krukenbergin tuumori). Aikana etäpesäkekasvainten evoluution, useita tunnettuja ja ehdokas syöpä kuljettaja tapahtumia rajattuja kasvaimen kehittyminen ja geneettiset eroavaisuudet etäpesäke primaarikasvaimen.
Selvitimme jos ehdokas kuljettajia tästä metastasointiin riittivät lisääntymään hajanainen mahasyöpä. Meidän syöpä mallintamismenetelmään käyttää in vitro
mahalaukun organoids ja mahdollistaa yhden insinööri geneettisen kuljettaja yhteydessä näiden syöpien ja tutkia prosessi metastaattisen evoluution ja onkogeenista polku eroavaisuuksia. Integrointi geneettinen analyysi ja biologisten mallinnus, päätimme itsenäinen rooli TGFBR2
(transformoiva kasvutekijä-β reseptori 2) synnyssä hajanainen mahasyövän. Meidän kokeellinen syövän mallinnus perustuu ilma-neste rajapinta ensisijaisen hiiren suolen kulttuuri, joka sisältää sekä epiteelin ja mesenkymaaliset elementtejä, tarkasti ssä toistetaan pitkäaikainen lisääntymistä, multilineage erilaistuminen, Wnt /Notch-riippuvaisen kantasolujen kapealla, ja peristaltiikkaa [11]. Me raportoitu analogista ensisijaisen mahalaukun organoidikappaleen kulttuuri, joka tarkasti ssä toistetaan multilineage epiteelin erilaistumista ja peruskudoselementeistä [12]. Viime aikoina olemme saavuttaneet vankan in vitro
onkogeenisia muutosta ensisijainen mahan, paksusuolen ja haiman organoids kautta mutaatiot Kras
ja Trp53
, jotka aiheuttavat dysplasiaan ja invaasiota in vitro
adenokarsinooma upon ihonalainen siirrettäväksi hiiriin [13]. Osoitamme toiminnallinen validointi ehdokas mahasyövän etäpesäke kuljettajia syöpään genomisesta profiloinnin tutkimuksia keskittyen mallintamiseen TGFBR2
kuljettaja todisteena periaatteen.
Tulokset
Diffuusi mahasyöpä ja metastasointiin
vuotiaana ja 37 vuotta, indeksi potilas (525) oli diagnosoitu vaiheen III (T3N1M0) huonosti eriytetty diffuusi mahalaukun adenokarsinooman (kuvio 1). Hänen 42-vuotias siskonsa todettiin diffuusi mahalaukun adenokarsinooman 2 kuukautta aikaisemmin. Perustuu suvussa mahasyövän ja epätavallisen nuorena puhkeamista, hän koki ituradan CDH1
mutaatio testaus. Potilas ja hänen sisarensa havaittiin ituradan silmukointikohtamutaatio intronissa 10 (c.1565 + 2insT). Tämä ituradan mutaatio myöhemmin raportoitu toisen perheen kanssa perinnöllinen diffuusi mahalaukun syöpä (HDGC) [14]. Potilas koki yhteensä gastrektomia poistaa hänen primaarikasvaimen ja havaittiin yksi imusolmuke etäpesäke. Hän sai standardin adjuvanttihoitona lukien yhdistetty kemoterapiaa (sisplatiini ja 5-fluorourasiilin) ja säteily. Kolmen vuoden kuluttua hänen ensimmäisellä kerralla, potilas kertoi progressiivinen alavatsan täyteläisyyttä. Tietokonetomografia (TT) osoitti suuri lantion massa vastaa vasemman munasarjojen etäpesäke (kuvio 1). Tämän jälkeen potilas koki laparotomy kahdenvälisillä salpingo munasarjan poisto ja biopsia lantion massa. Patologiset tutkimukset osoittivat, metastaattinen adenokarsinooma, joihin munasarja, muuten kutsutaan krukenbergin tuumori, samalla histologinen ulkonäkö kuin primaarikasvaimen. Eräs tutkimus kertoo, että joukossa hajanainen mahasyövän metastasoitunutta levittämiseen, munasarja oli metastaattinen sivuston 28,8% tapauksista [15]. Siten munasarja on yhteinen sivusto etäpesäkkeitä.
Cancer Genomikartoituksen analyysi
Sekä exome ja koko genomin pariksi-end sekvensointi tehtiin primaarikasvaimen, munasarjojen etäpesäkkeitä, ja normaali kudos, joka sisälsi veren ja normaali maha- kudoksen (Additional tiedosto 1: Taulukko S1). Kudosta imusolmuke etäpesäke ei ollut saatavilla analysointia varten. Useita sekvensointi menetelmiä käytettiin korvaamaan siinä määrin normaalien strooman seoksen, joka on suoraa seurausta infiltroiva invasiivisuuden hajanaista mahasyövän alatyyppi. Selvitimme laajuus normaalin genomin seoksen ja korjataan sisällyttäminen normaalin DNA (Additional tiedosto 1: Methods). Monimutkaisuuden vuoksi kasvain näytteet, suoritimme ylimääräisen kierroksen kohdennettuja sekvensoinnin vahvistaa läsnäoloa mutaatioiden ja muita geneettisiä poikkeamia, joka tapahtui eksonit, lähellä eksoni rajojen tai promoottorit.
Kaiken kaikkiaan saimme yli 100 × keskimääräinen kattavuus kunkin exome ja yleensä luottaneet exome tietojen löytämisen koodausalueen mutaatioita. Sillä koko genomin sekvensointi, meillä oli yli 60 × keskimääräinen kattavuus ensisijaisen syövän koko genomin näyte ja 30 × varten metastaattisen genomin. Koko Genomikartoituksen käytettiin tunnistamaan suuremman mittakaavan geneettisten poikkeavuuksien kuten kopioluvun vaihtelu (CNVs), alleelinen epätasapaino, järjestelyihin sekä muunkinlaisia, järjestelyihin. Sen jälkeen kohdistus teimme variantti soittamalla tunnistaa somaattisten mutaatioiden ja muihin luokkiin geneettisten poikkeavuuksien. Tämä sisältyi somaattiset mutaatiot, lisäys-poistot (indeleitä), CNVs, keskeytys- Heterotsygoottisuuden alueiden (LOH), ja syöpä uudelleenjärjestelyt (Additional tiedosto 1: Taulukko S3 ja taulukko S4). Kontrollina yhden nukleotidin muunnos vaatii, me genotyyppi näytteet Affymetrix 6,0 yhden emäksen monimuotoisuus (SNP) matriisia; vertasimme genotyypit tunnistettuihin SNP sekvenssistä tiedot. Konkordanssi of exome ja koko genomin SNP dataa array data oli 99%.
Koodausalueen mutaatioita ja validointi syvä sekvensoimalla
Olemme tunnistettu mutaatioita, jotka tapahtuivat eksonit ja introni mutaatioita 100 emästä eksonin rajan ja tulokset on koottu Additional tiedosto 1: Taulukko S2. Kuten edellä on todettu, kasvain näytteet oli monimutkainen koostumus, joka vähensi sekvenssi kattaa joitakin mutaatioita. Meillä eteni ylimääräinen kierros kohdennettujen sekvensoinnin vahvistaa nämä mutaatiot ja määrittää niiden läsnäolo molemmissa kasvaimia. Suunnittelimme määritystä syvälle kohdennettuja resequencing joka peitti noin 300 perustaa ympäri tietyn mutaation lokuksen (Additional tiedosto 1: Taulukko S5). Keskimääräinen suunnattu sekvensointi kattavuus jokaisen otaksuttu mutaation tai loci oli 278 × normaalia, 251 × varten primaarikasvaimen ja 152 × varten etäpesäkkeitä.
Näiden kahden kasvaimia, me itsenäisesti validoitu yhteensä 77 mutaatioita, jotka tapahtuivat tai proksimaalisesti eksonit (Additional tiedosto 1: menetelmät ja taulukko S5). Validoitu geneettinen poikkeamia sisältyvät: (1) ei-synonyymi mutaatioita, (2) synonyymi mutaatioita, (3) insertioita, tai (4) poistot. Kohdistettuun sekvensointi tietoja, päätimme mutaatio alleelifrekvenssi (MAF) välillä primaarikasvaimen ja etäpesäkkeiden kunkin mutaation. Tämä edellyttää määrittämisen osa sekvenssin luetaan mutaation verrattuna referenssisekvenssiin lukee. Pystyimme tunnistamaan, mitkä mutaatiot olivat yhteisiä tai yksinoikeudella primaarikasvaimen vs. etäpesäke. Niistä 77 validoitu mutaatiot, jakelua oli sellainen, että mutaatiot olivat yleensä ainutlaatuisia joko primaarikasvaimen tai metastaattinen päällä. Esimerkiksi, primaarikasvaimen oli kahdeksan mutaatioita, jotka eivät olleet läsnä etäpesäkkeitä, kun etäpesäkkeitä oli 37 mutaatioita ei ole läsnä primaarikasvaimen. Yhteinen sekä syövät olivat 32 mutaatioita.
Koska kolmen vuoden välein ennen havaitsemista etäpesäkkeiden, on olemassa mahdollisuus, että metastaasin mutaatioiden tapahtui riippumatta primaarikasvaimen. Mutaatiot ominaisia primaarikasvaimen, jotka eivät olleet läsnä munasarjojen etäpesäkkeitä saattanut olla seurausta satunnainen geneettisen drift. Mutaatiot yhteisiä sekä osoittaa yhteinen alkuperä mutta tarkka ajoitus eriyttäminen kahden kasvaimia on vähemmän selkeä kuin huomata ne mutaatiot pienemmillä MAF. Osaa näistä geeneistä oli korkea MAF arvot, mikä osoittaa suurempi todennäköisyys olla läsnä kaikissa klonaalisia populaatioita primaarikasvaimen tai etäpesäke. Kuten kuvataan myöhemmin, nämä geenit priorisoitiin edelleen testauksiin mahalaukun organoids.
Mutaatiot vaikuttavat geenien toiminnan
joukossa mutaatioita, jotka olivat ulkoisesti validoitu, olemme keskittyneet osajoukko mutaatioista johtavat aminohapposubstituutioihin, ennenaikainen pysäytys kodoneja ja indeleitä että muuttaneet avoin lukukehys. Myöhemmin määritimme jos nämä koodaus mutaatiot olivat mahdollisesti haitallisia geenin toiminnan käyttämällä useita Ennustusalgoritmien kuten Polyphen [16] ja SIFT [17] mm. Perustuen MAF tiedot kunkin mutaation, päätimme ovatko nämä mutaatiot jossa mahdollinen vahingollinen vaikutus geenin tuotteet olivat yhteisiä tai yksinoikeudella primaarikasvaimen ja etäpesäkkeiden (kuva 2). Kuva 2 vertailu geneettisen poikkeamia primaarikasvaimen ja etäpesäke. Yhteiset vs.
yksinomainen geneettisiä poikkeavuuksia verrataan kahden kasvaimen genomeja. (A) Genes koodausvälineillä mutaatioita, joilla on mahdollisesti haitallisia vaikutuksia on lueteltu. Nämä geenit luokitellaan sen mukaan, onko ne eivät sisällä (punainen merkkiä) tai yleinen (vihreä merkkiä) primaarikasvaimen ja etäpesäke. Mutaatiot kaikki johtavat muutoksiin aminohappokoostumus geenituotteen ja havaittiin olevan merkittävää muutosta, jolla on suuri todennäköisyys vaikuttaa geenituotteen toimintaa. (B) yhteenveto kromosomipoikkeavuuksien näkyy koko syövän genomin molemmat kasvaimia. Tämä sisältää kopioluvun vaihtelu (CNV) tai Heterotsygotian menetys (LOH). Punainen lohkot ilmaisee tapahtumien yksinoikeudella primaarikasvaimen tai etäpesäke. Vihreä lohkot osoittavat tapahtumia yhteisiä molemmille. Tapahtumien määrä per kromosomi on lueteltu kunkin lohkon. Nuolet osoittavat LOH tapahtumia tai deleetioita, jotka kattavat p varsi, q käsivarren tai koko kromosomi. Punaiset nuolet osoittavat kromosomimuutokset jotka ovat yksinomaan ja vihreät nuolet osoittavat tapahtumia, jotka ovat yhteisiä.
Sen osajoukko vahingollisia mutaatioita, teimme biologisiksi koulutusjakson analyysi, kirjallisuuskatsaus ja vertaaminen Cancer Genome Atlas tietoja saatavilla hajanainen mahasyövän. Tämä tunnistaa joukon tunnettuja syövän geenien ja todennäköisesti syöpään liittyvien ehdokkaita mutaatioita, jotka todennäköisesti ollut vaikutusta proteiinin toimintaan. Keskityimme useita Hakijan geenien (taulukko 1), joka oli aiemmin osoitettu olevan onkogeenisiä tai tiedettiin tuumorisuppressoreilla bialleelisilla muutoksia läsnä syöpä genomes.Table 1 Syöpä onkogeenien kanssa lisäyksiä tai syövän kuljettajien bialleelisilla tapahtumia
Alkuperäinen
Todettu tai ehdokas syöpä kuljettaja
Biallelic tapahtuma
alleelisia muutos 1
mutaatio tai genomista poikkeamia
Chr
Chr asema tai väli
alleelisia muutos 2
Unique ensisijaiseen
FGFR2 *
Amplification
6-kertainen vahvistus
10
117820033-119748751
yhteisen ensisijaisen kasvaimen ja etäpesäkkeiden
CDH1
Kyllä
poistaminen
osittainen poistaminen eksonin 9
16
68847326-68847403
Ituradan mutaatio CDH1
TP53
Kyllä
5 'silmukointikohtamutaatio
Poikkeava liitos
17
7578370
Hemizygous menetys 17p arm
Ainutlaatuinen etäpesäke
TGFBR2
Kyllä
Lukukehyksensiirtomutageenit Indel
Stop-kodonin eksonin 4
3
30691871
Hemizygous poistetaan villityypin TGFBR2
locus
PCDH7
Kyllä
Missense
S87R
4
30723305
Hemizygous poistetaan villityypin 4 varren
FERMT1
loci
Kyllä
Loss heterotsygoottisuuden
FERMT
1 sijaitsee 20p12.3
20
FERMT
1 mutaatio
BMP7
loci
Kyllä
menettäminen heterotsygoottisuuden
BMP7
sijaitsee 20q13.3
20
BMP7
mutaatio
Chr: kromosomi.
Kopioi numero muunnelmia ja alleeliset epätasapainot erottaa ensisijaisen kasvaimen etäpesäke
Totesimme laajemmassa mittakaavassa genomista poikkeavuuksia että eriytetty ensisijainen päässä etäpesäke (kuviot 2b ja 3a). Tämä sisältyi kopiomäärä muutoksia ja LOH tapahtumia. Ainutlaatuinen primaarikasvaimen oli kaksi genomista monistuksissa kromosomeissa 5 ja 10, ja kaksi käännellen kromosomeissa 15 ja 16 (Additional tiedosto 1: Taulukko S4). Kromosomissa 10 vahvistus kattoi 1,66 Mb välein. Harkittaessa poistojen tai alleeliset epätasapainot, ainoa merkittävä tapahtuma, joka merkittiin mukana menetys p varren kromosomin 17. Kuva 3 geneettinen divergenssi munasarjojen etäpesäke ensisijaisesta mahasyövän kriittisiin ehdokas kuljettajia. Genominen mutaation asemassa, kopioiden määrä variaatioita (CNV) alueita tai menettämisestä Heterotsygoottisuuden (LOH) välein näkyvät päässä syöpään genomeja. Sillä kromosomi tontteja, Y-akseli tarkoittaa asemaa vastaavaan kromosomi, sen pituus megabases (MB) ja ideogrammi nimitys näkyy vasemmalla kopiomäärä profiilin. Vahingollisia mutaatioita näkyvät boxed nuolia geenin symboli. (A) genomin jaetaan laajasti syövän erityisiä CNVs ja LOH välein on koottu kaikkien kromosomien primaarikasvaimen ja etäpesäkkeiden. (B) kromosomissa 3 etäpesäkkeitä oli ainutlaatuinen bialleelisen tapahtumia, joihin liittyy haitallisia TGFBR2
mutaation ja genominen deleetio vaikuttaa muiden alleeli kuten nähdään selvimmin LOH välein. Toissijainen genomisen deleetioita, LOH on osoitettava muutos vähäinen alleelifrekvenssi suhdearvo -1 ja korreloi genomisen deleetion. (C) kromosomissa 10 tarkoitetaan FGFR2
geeni sijaitsee genomivahvistuksen alue näkyy ainoastaan ensisijaisen eikä etäpesäkkeitä. Vahvistus on huomattava, punainen ympyrä.
Toisin primaarikasvaimen, etäpesäketuumorikudoksen oli lukuisia kromosomaalisen mittakaavassa LOH tapahtumia ja genomisen deleetiot vaikuttavat 12 eri kromosomeja, joista suurin osa oli ainutlaatuinen metastaattinen kasvain (kuvio 2) . Tämä sisältyi useita deleetioita ja kopioi neutraali LOH tapahtumia, jotka yksityiskohtaisesti Additional tiedosto 1: Taulukko S3. Oli viisinkertaisesti genomivahvistuksen kromosomissa 2 mutta mitään erityisiä tunnettuja geenejä olemassa sairaaseen välein. Ei ollut havaittavissa välistä kromosomitranslokaation joko primaarikasvaimen tai etäpesäke genomeja. Muita syövän uudelleenjärjestelyjä tunnistettiin mutta eivät viitanneet poikkeamia millään Hakijan geeneistä (Additional tiedosto 1: Taulukko S4). Oli viitteitä suuren mittakaavan perimän epävakaisuuden perusteella alleeliset epätasapaino analyysi; Kromosomit 14, 17, 20, ja 22 kaikki mukana koko kromosomi.
varten kopiomäärä poikkeavuuksien alleeliset epätasapainot, tunnistimme yksinomainen versus yhteisiä tapahtumia välillä primaarikasvaimen ja etäpesäke. Ainoa yhteinen geneettinen poikkeavuus mukana p käsivarteen kromosomin 17. Kaiken puute päällekkäisyys oli osoitus merkittävä geneettinen poikkeama primaarikasvaimen ja etäpesäkkeiden huolimatta sama alkuperä kuin merkitty yhteinen mutaatioita kriittisissä tuumorisuppressoreilla.
Genomista välein ja LOH, kopioiden määrä poikkeavuus ja uudelleenjärjestely tapahtumia verrattiin asemaan validoitu geenimutaatioita. Tämä integroitu analyysi mainitsi useita geenejä, jotka oli kaksialleelisten muutokset ovat mukana sekä menetys villityypin alleelin suuresta väli genomista poikkeavuus ja mutantti alleeli. Tulokset geenien bialleelisilla osumia katsottiin vahvaa ehdokasta menettämisestä toiminnon osallistuminen syövän (taulukko 1).
Tunnistaminen syövän kuljettajien yhteisiä primaarikasvaimen ja etäpesäkkeiden
Sekä ensisijainen ja etäpesäkkeiden sisältyvät syöpä kuljettajan tapahtumia, jotka saattoivat olla kriittinen kasvaimien synnyn yhteydessä alkuperäisen CDH1
mutaatio (taulukko 1, kuvio 2). Sen lisäksi, että ituradan CDH1
introni mutaatio, toinen CDH1
alleeli oli somaattisten 77 emäsparin genomisen poistamalla osa eksonin 9, joka vaikuttaa alavirran koodaavien alueiden samoin. CDH1
somaattinen mutaatio oli sama molemmissa ensisijainen ja metastaattinen mahasyövän genomeja, mikä osoittaa yhteinen geneettinen alkuperä ja tarjoamalla vahvaa geneettisiä todisteita siitä, että tämä kuljettaja oli ratkaiseva merkitys diffuusi mahan kasvainten synnyssä. Vaikuttavia mutaatioita CDH1
eksonin 9, jotka johtavat menetykseen proteiinin ilmentymisen on usein havaittu hajanainen mahasyövässä [18] - [20]. Tämä eksonin: n aminohapposekvenssi on oletettu kalsiumia sitovaan kohtaan, joka on todennäköisesti tärkeä reseptorin toimintaa.
Primaarinen ja metastaattinen kasvaimen myös jaettu bialleelisen silmukointidonoripaikan mutaatio (c.559 + 1G > A) viidennen intronin TP53
ja kromosomissa 17p Loh tapahtuma kattaa TP53
lokuksen (Additional tiedosto 1: Kuva S1). TP53
liitos mutaatio keskeyttää Silmukointi [21] ja on aiemmin raportoitu syöpä mutaatio [22], [23]. Analyysit satunnaista ja perinnöllinen syöpien ovat tunnistaneet TP53
mutaatioita, jotka tapahtuvat samanaikaisesti CDH1
mutaatio [24], [25]. CDH1
inaktivaatio maha- parietaalisoluista ei aiheuta mahalaukun karsinooma, viittaa siihen, että menetys CDH1
on riittämätön kasvaimen aloittamista [26]. Kuitenkin kaksinkertainen ehdollinen tyrmäys CDH1
ja TP53
indusoi kehittymistä hajanainen mahakarsinooman [26]. Mielenkiintoista on, että genominen väli LOH tapahtumaa, joka vaikuttaa TP53
lokuksessa oli suurempi etäpesäkkeitä verrattuna primaarikasvaimen. Tämä saattaa johtua siitä, että riippumattomien perimän epävakaisuuden tapahtumista annetaan vahva valinta bialleelisten menetys TP53 funktion.
FGFRis kannekelpoinen syöpä kuljettaja yksinomainen ensisijaiseen mahakasvaimen
primaarikasvaimen, oli kuusinkertainen genomista alueen monistamiseen kromosomin 10 q käsivarteen ja katettu väli 1,66 Mb. Tässä genomialuetta oli onkogeenisen Hakijan FGFR2
myös nimitystä fibroblastikasvutekijäreseptori 2 (kuvio 3c). Tämä vahvistettiin useita menetelmiä, kuten sekvensointi, erilaisia analyysi ja validointi kvantitatiivinen PCR. FGFR2 on transmembraaninen reseptori, joka toimii osana avaimen signaalinvälitysreitin säätelemällä kudosten korjaamiseen ja sikiön kehityksen joukossa monia muita toimintoja [26].
Validoimiseksi esiintyvyys FGFR2
vahvistus diffuuseihin verrattuna suoliston mahasyövistä kartoitettiin 37 hajanainen ja 27 suoliston alatyyppi ensisijainen mahakasvaimen näytteiden digitaalinen PCR [27]. Aikaisemmin osoitimme, että tämä menetelmä on täysin herkkä havaitsemaan kopiomäärä poikkeavuus jopa yhteydessä normaalin diploid DNA laimentavaa kasvain DNA. Tutkimuksemme osoittaa, FGFR2
vahvistus neljä 37 (11%) diffuusi kasvain näytteet, jotka oli poissa suoliston alatyypin näytteet (kuvio 4a). Kuva 4 esiintyvyys FGFR2 ihmisen mahalaukun kasvaimet ja sen osuus soluproliferaatioon. (A) Yksittäisiä mahasyöpä näytteet arvioitiin kvantitatiivinen digitaalinen PCR määrittää FGFR2 genomista kopiomäärä. Mustat pisteet ovat hajanainen syöpien. Punaisia pisteitä osoittavat suoliston alatyypin mahasyövän. (B) geneettiset ominaisuudet AGS (FGFR2
diploidi) ja KatoIII (FGFR2 monistettu) mahasyöpä solulinjat on esitetty. (C) Prosenttia selviytymisen AGS syövän solulinja näytetään FGFR2 estäjiä vaihtelevia spesifisyys. (D) KatoIII hajanainen mahasyöpä solulinja käsiteltiin FGFR2 estäjiä vaihtelevia spesifisyys. Y-akseli kuvaa eloonjäämisprosentteina vastaan of the X-akselilla log pitoisuuksilla. Kaikissa paneelit, virhepylväät edustavat keskivirhe keskiarvon. Ero prosentteina solujen eloonjäämisen välillä KatoIII ja AGS solut oli tilastollisesti merkitsevä (P
< 0,05) kolmessa korkeimmat pitoisuudet kaikkien huumeiden, paitsi Brivanib joka oli vain merkitsevä korkein pitoisuus.
Tueksi rooli hakijan, FGFR2
vahvistus on läsnä useissa mahalaukun syövän solulinjat [28], [29] ja sen jälkeen on raportoitu eri maha-suolikanavan pahanlaatuisten kasvainten, kuten ruokatorven adenokarsinooma [30]. Lisäksi syövän solulinjojen FGFR2-spesifinen pienmolekyylisalpaajien tai shRNAs johtaa voimakas kasvun estäminen [28] viittaa siihen, toiminnallinen rooli FGFR2
monistusta diffuusi alatyyppi.
Toiminnallinen analyysi FGFR2 kuljettajan yhdessä CDH1 ja TP53
Havaitsimme kaksi esimerkkiä ensisijaisena hajanainen mahasyövän kanssa yhteistyössä esiintyminen tunnettujen ja oletettua syöpä ajurit mukana CDH1
, TP53
, ja FGFR2
nähty indeksin potilaalla . Ensimmäinen esimerkki sisältyi hajanainen mahasyövän näyte, joka oli yksi mahalaukun adenokarsinooman analysoitiin TCGA. Käyttämällä cBio TCGA portaalin [10], tunnistimme potilaan (TCGA-BR-6803), joka oli samanlainen täydentää geneettisiä poikkeavuuksia CDH1
, TP53
, ja FGFR2
, jotka kaikki ovat olleet aikaisemmin kuvattu syövän nähty COSMIC syövän mutaatio arkistoon. Tämä sisälsi seuraavat: missensemutaatio vuonna CDH1
(D254Y), joka on kuvattu kolmessa muiden syöpien; missensemutaatio (L130F) vuonna TP53
jossa mutaatiot tässä kodonissa on raportoitu 37 muiden syöpien; FGFR2
vahvistus, joka me ja muut ovat tunnistettu diffuusi mahasyövässä.
Toisena esimerkkinä, olemme tunnistaneet ihmisen diffundoitua mahasyövän solulinja, KatoIII, joka on samanlainen koostumus, geneettisten poikkeavuuksien vaikuttavat samalla syöpägeenit ensisijaisena kasvain meidän indeksin potilaan. KatoIII on CDH1
mutaatio johtaa introni lisäämästä mRNA [31], [32], joka on TP53
mutaatio johtaa täydellisen geenin poisto [33] ja FGFR2
vahvistus [29] (kuvio 4b). Tämä solulinja annettiin voimme arvioida mahdolliset onkogeeniset roolia FGFR2
monistusta geneettisen yhteydessä CDH1
ja TP53
mutaatioita, samanlainen indeksin potilaan primaarikasvaimen.
Määrittämiseksi panos ja FGFR signaloinnin neoplastisen kasvun, käsittelimme KatoIII solut, joissa on useita FGFR2 pienimolekyylisiä tyrosiinikinaasi-estäjät (TKI), mukaan lukien Brivanib, TKI258, Ponatinib, ja AZD4547 [34]. Kontrollina käytimme mahasyövän solulinja AGS, joka on villityypin ja FGFR2
, CDH1
, ja TP53
, mutta on mutaatiot KRAS
ja PIK3CA
[35] ( kuva 4b). Kaikki FGFR2 estäjät indusoituvaa solukuolemaa KatoIII mutta ei AGS-soluja (kuvio 4c ja d). Voimakkaimmat näistä TKI, AZD4547, on IC 50 noin 2 nM KatoIII soluissa ja 39580 nM AGS-soluissa (kuvio 4c ja d). Kukin estäjien osoitti tilastollisesti merkitsevästi vähemmän IC 50 FGFR2-monistettiin KatoIII soluja verrattuna ei-FGFR2-monistettiin AGS solujen yhdelläkään testatulla pitoisuudella (kuvio 4c ja d).
Sitä vastoin hoito KatoIII ja AGS solujen sytotoksisten kemoterapeuttisten aineiden kuten paklitakselin, 5-fluorourasiili ja karboplatiini ei ollut merkittävää vaikutusta joko KatoIII tai AGS linjat, joilla on samanlaiset IC 50 yksilöity kussakin (Additional tiedosto 1: Taulukko S7). Sillä AZD4547, 20000-kertainen ero herkkyydessä FGFR estäjiä viittaa siihen, että FGF signalointi on kriittinen kuljettaja CDH1
-initiated mahalaukun solujen lisääntymistä ja tämä TKI on potentiaalinen täsmähoitoihin heikossa alatyypin syövissä kätkeminen FGFR2
monistuksissa.
Kaksialleeliset inaktivointi TGFBRis yksinoikeudella munasarjojen etäpesäkkeiden
Genetic eroavuus oli ilmeinen;
 Tutkijat manipuloivat suoliston bakteerilajeja ruokavalion avulla
Tutkijat manipuloivat suoliston bakteerilajeja ruokavalion avulla
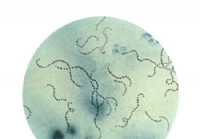 Mikä on streptokokki-infektio?
Mikä on streptokokki-infektio?
 Hepatiitti
Hepatiitti
 Kahvi auttaa kehittämään suoliston terveitä mikrobeja ja tukkii suoliston liikkeitä
Kahvi auttaa kehittämään suoliston terveitä mikrobeja ja tukkii suoliston liikkeitä
 Kuinka kauan kestää, että lapsi toipuu umpilisäkkeen poistoleikkauksesta?
Kuinka kauan kestää, että lapsi toipuu umpilisäkkeen poistoleikkauksesta?
 Mikä on pitkittäinen haima-ojejunostomia?
Mikä on pitkittäinen haima-ojejunostomia?
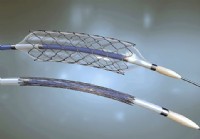 Kuinka pitkäksi aikaa sappistenttiä voi jättää?
Mikä on sappistentti? Sappistentti on putki, joka on sijoitettu kirurgisesti pitämään sappitiehyen auki. Sappistentti on muovi- tai metalliputki, joka on asetettu sappitiehyen. Sappien stentointi
Kuinka pitkäksi aikaa sappistenttiä voi jättää?
Mikä on sappistentti? Sappistentti on putki, joka on sijoitettu kirurgisesti pitämään sappitiehyen auki. Sappistentti on muovi- tai metalliputki, joka on asetettu sappitiehyen. Sappien stentointi
 Välimeren ruokavalio edistää tervettä ikääntymistä terveemmällä suoliston mikrobiomilla
Uusi tutkimus julkaistiin verkossa lehdessä Gut raportoi helmikuussa 2020 hämmästyttäviä terveyttä edistäviä vaikutuksia, kun siirtyminen Välimeren ruokavalioon vain vuodeksi. Tulokset johtuivat ter
Välimeren ruokavalio edistää tervettä ikääntymistä terveemmällä suoliston mikrobiomilla
Uusi tutkimus julkaistiin verkossa lehdessä Gut raportoi helmikuussa 2020 hämmästyttäviä terveyttä edistäviä vaikutuksia, kun siirtyminen Välimeren ruokavalioon vain vuodeksi. Tulokset johtuivat ter
 Terveysvinkki:merkit, jotka voivat varoittaa laktoosi-intoleranssista
Viimeisimmät Digestion-uutiset Kun munuaissiirto epäonnistuu, on parasta yrittää uudelleen Älä anna närästyksen pilata lomajuhlasi Maksansiirtojen kysyntä kasvaa jyrkästi Pieniannoksiset TT-skannauks
Terveysvinkki:merkit, jotka voivat varoittaa laktoosi-intoleranssista
Viimeisimmät Digestion-uutiset Kun munuaissiirto epäonnistuu, on parasta yrittää uudelleen Älä anna närästyksen pilata lomajuhlasi Maksansiirtojen kysyntä kasvaa jyrkästi Pieniannoksiset TT-skannauks