absztrakt katalógusa
Háttér katalógusa
Fertőzés Helicobacter pylori katalógusa kiváltja krónikus gyomor- gyulladás fejlődhet sorvadás és gyomor adenokarcinóma. Polarizációja makrofágok jellemző mind a rák és a fertőzés, és elősegítheti progresszió vagy felbontása betegség. Azonban a szerepe a makrofágok és a polarizációs során H. pylori fertőzés katalógusa nem jól definiált. katalógusa Módszertan /fő eredményei katalógusa használatával egér modellben a fertőzés és a gyomor biopszia 29 magánszemélyek elemeztük makrofág toborzás és a polarizáció során H. pylori Következtetések /Jelentősége katalógusa Ezek az eredmények azt mutatják, hogy az oltás egerek ellen H. pylori Citation: Quiding-Järbrink M, Raghavan S, M Sundquist (2010) Fokozott M1 makrofág polarizáció Humán Helicobacter pylori katalógusa -asszociált sorvadásos gyomorhurut és A beoltott egerek. PLoS ONE 5 (11): e15018. doi: 10,1371 /journal.pone.0015018 katalógusa Szerkesztő: Niyaz Ahmed, University of Hyderabad, India katalógusa Beérkezett: augusztus 26, 2010; Elfogadva: október 7, 2010; Megjelent: november 23, 2010 katalógusa Copyright © 2010 Quiding-Järbrink et al. Ez egy nyílt hozzáférésű cikk feltételei szerint terjeszthető a Creative Commons Nevezd meg! Licenc, amely engedélyezi a korlátlan használatát, a forgalmazás és a reprodukció bármilyen adathordozón, feltéve, hogy az eredeti szerző és a forrás jóváírásra. Katalógusa Forrás: Ez a tanulmány támogatta a kiválósági központ MIVAC által finanszírozott svéd stratégiai Kutatási Alapítvány, a Adlerbert Research Foundation, Wilhelm & Martina Lundgren alapítványa, Inga-Britt & Arne Lundberg alapítása, és az Sahlgrenska Egyetemi Kórház. A finanszírozók nem volt szerepe a tanulmány tervezés, adatgyűjtés és elemzés, döntés, hogy közzéteszi, vagy a készítmény a kézirat. Katalógusa Érdekütközés: A szerzők kijelentették, hogy nem ellentétes érdekek léteznek. Katalógusa Bevezető Helicobacter pylori Egy erős pro-gyulladásos választ társul megnövekedett reaktív oxigén és nitrogén gyökök a gyomornyálkahártya [3], amelyek elősegítik a rák fejlődését. [4] Például fertőzött egerek H. pylori katalógusa hat hónapig, nagyobb gyakorisággal a gyomorsav mutációk képest fertőzött egerekben [5]. Ezen túlmenően, az egerek, amelyek hiányosak az enzim indukálható nitrogén-oxid-szintáz (iNOS) van egy csökkentett incidenciája gyomorrák után H. pylori M1 makrofágok jellemzően részt venni a kezdeti immunválasz a behatoló mikroorganizmusok és elősegíti a T helper (Th) 1 immunitás, míg az M2 makrofágok során indukálódik a felbontás fázis gyulladás és részt vesznek a törmelék öblítő olyan lágy szövetek, és előmozdítása Th2 immunitás [8], [9]. Polarizációja makrofágok által irányított mikrokörnyezet. M1 makrofágok által indukált interferon-γ és mikrobiális termékek, mint a lipopoliszacharid [9]. Másrészt, az M2 makrofágok által indukált TH2- vagy gyulladásellenes citokinek és növekedési faktorok, beleértve az IL-4, IL-10 és a transzformáló növekedési faktor-β [8], [9]. alatt H. pylori Megnövekedett gyakoriságát makrofágok, eozinofilek és neutrofilek a gyomornyálkahártya után H. pylori fertőzés katalógusa katalógusa A felvételi veleszületett sejteket a fertőzés helyén előfeltétele fertőző ellenőrzés. Nem csak a veleszületett sejtek, például makrofágok és neutrofilek, részt baktériumölésben; ők is termelnek a gyulladásos mediátorok, amely előkészítette a terepet a következő immunválaszt. Annak vizsgálatára, a felhalmozódása a veleszületett sejtek a gyomor nyálkahártya alatt H. pylori Az eozinofil sejteket definiált CD11b + Siglec-F + sejtek (ábra. 1C, [17]). Ezek a sejtek kifejezett köztes szintjeit F4 /80 volt, és magas Side Scatter profil amikor áramlási citometriával analizáltuk (ábra. 1E). Krezilibolya festése rendezve CD11b + Siglec-F + sejtek megerősítette eosinophil morfológia (ábra. 1C). A frekvencia az eozinofil sejtek a gyomor nyálkahártyájának megduplázódott nyolc hét után, és tovább növekedett 26 hét után a fertőzés (ábra. 1F). Katalógusa neutrofil sejteket definiált CD11b + Gr1 + sejtek (ábra . 1D). Mivel a Gr1 antitest felismeri mind a Ly6C és Ly6G epitóp azt megerősítette, hogy minden CD11b + Gr1 + sejtek kifejezett specifikus neutrofil marker Ly6G (ábra. 1D és E). A frekvencia a neutrofilek megnövekedett 10-szeres nyolc héttel a fertőzés után, és tovább növekedett 26 hét után (ábra. 1F). Így alatt H. pylori fertőzés katalógusa van egy szekvenciális neutrofilek felhalmozódását és eozinofil, majd makrofágok a gyomor lamina propria. katalógusa jellemzése gyomor dendritikus katalógusa jellemzésére gyomor DC, először azonosították a lakosság CD11c + MHC-II + sejtek (2A.). Amikor ezeket a sejteket tovább analizáltuk expressziójának F4 /80 és a αE integrin lánc CD103, a fele a CD11c + MHC-II + sejteket azonosították F4 /80 + makrofágok (2A. ). Azonban, többek között a CD11c + MHC-II + sejtek hogy hiányzott expressziója F4 /80, két populáció feltételezett dendritikus differenciális expressziója CD103 lehet megkülönböztetni (2A.). Mivel a sok fluorokrómok szükséges úgy döntöttünk, hogy csak jellemzik a gyomor CD103 + Gyomor makrofágok és CD103 + DS nem upregulate kostimulációs molekulák után H. pylori fertőzés katalógusa katalógusa Annak érdekében, hogy vizsgálja meg a lehetséges hatásait H. pylori M1 polarizációja gyomor makrofágok során H. pylori fertőzés katalógusa katalógusa Mivel eredményeink azt javasolta, hogy a gyomor makrofágok esetleg nem lesz teljesen aktiválva során H. pylori fertőzés katalógusa, mi jellemzi ezeket a sejteket tovább vizsgálta a M1 /M2 polarizáció. Annak megállapításához, makrofág polarizáció során H. pylori forrásának azonosítása iNOS és CXCL11 a gyomornyálkahártya, a makrofágok (CD11b + Gr1 -Siglec-F -MHC-II +), eozinofil (CD11b + Gr1 -Siglec-F + MHC-II -), és a maradék sejteket után kapuzássai ki makrofágok és eozinofil (CD11b - és CD11b + Gr1 +) soroltam poolozott gyomor lamina propria sejtek fertőzött egerek H. pylori elleni védőoltás H. pylori katalógusa erősíti makrofág M1 polarizáció katalógusa Védő elleni immunizálás H. pylori katalógusa általában társított gyors fejlődés a gyomor Th1 [19]. Mivel a Th1 citokin interferon-γ indukál makrofág M1 polarizáció [9] azt vizsgáltuk, hogy a vakcinázás befolyásolhatja makrofág polarizáció során H. pylori fertőzés Ezen kívül, elemeztük a frekvencia a makrofágok a gyomor nyálkahártyáján az immunizálás és egerek áramlási citometriával. Relatív, hogy fertőzött-csak egerekben, immunizált egerek szignifikánsan nagyobb gyakorisággal jelentkeznek gyomor makrofágok három héttel a fertőzés után (ábra. 5C). Annak ellenére azonban, hogy a makrofágok vettek fel a gyomor nyálkahártyáján az immunizálás és egerekben a makrofágok nem upregulate expresszióját CD86 vagy az MHC-II viszonyítva fertőzött csak-egerek (ábra. 5D). Ezek az eredmények azt mutatják, hogy a sikeres vakcináció H. pylori dúsításának makrofág M1 polarizáció humán atrófiás gasztritisz következő betöltött szerepét vizsgáltuk a H. pylori fertőzés Annak vizsgálatára, hogy a fokozott mRNS expressziója M1 és M2 markerek is fordítja megnövekedett fehérje szinten, a teljes fehérjéket extraháltunk antrális biopsziás minták és a koncentrációt az iNOS és CCL18 ELISA-val meghatároztuk. A gyomor biopszia magánszemélyek sorvadásos gyomorhurut volt, csak a fehérje kivonással egy önkéntes, amelyet különbözőképpen jelzett 6B. Fele a H. pylori Mindent összevetve, ezek a megállapítások jelzik jelenléte mind az M1 és M2 makrofágok a gyomor nyálkahártyájának az H. pylori Ebben a tanulmányban megvizsgáltuk a polarizáció a gyomor makrofágok a krónikus H. pylori fertőzés Ezzel szemben a humán H . pylori Megfigyeltük szekvenciális felvételét veleszületett sejtek a gyomor nyálkahártya a SS1-fertőzött egerek, a makrofágok felhalmozódó meglehetősen késői során fertőzés (26 hét). Ezzel ellentétben, a frekvenciák a gyomor neutrofilek és eozinofileket a gyomornyálkahártya megnövekedett nyolc héttel a fertőzés után, és szintje emelkedett maradt 26 hét. A felhalmozódás makrofágok történt sokkal gyorsabb a vakcinált egerekben, amely esetben a gyakorisága gyomor makrofágok növeltük már három héttel a fertőzés után. Korábban, neutrofilek kimutatták, hogy kell felvenni, hogy a gyomor nyálkahártya a SS1-fertőzött egerek két szakaszban, egy korai fázisában tetőzik 1-2 nappal a fertőzés után, és egy késői fázisú kezdve 2-3 héttel a fertőzés után [24]. A toborzás minta, hasonló a neutrofilek, szintén leírtak gyomor makrofágok [24]. Azonban a makrofágok határozták meg CD11b + Gr1 - sejtek [24], a sejt populáció kezünkben elsősorban áll eozinofil (ld. 1. és 2. ábra). Asim et al. definiált makrofágok CD11b + F4 /80 + sejtek, és a megfigyelt egy korai csúcs makrofág számot a gyomornyálkahártya 1-2 nappal a fertőzés után a H. pylori tudtunk azonosítani két populáció dendritikus gyomornyálkahártya egerek. Mindkét alcsoport magas szinten expresszált CD11c és az MHC-II, hiányzott kifejezése a makrofág marker F4 /80, de különbözött tekintetében kifejezése CD103. A frekvencia a gyomor CD103 + DS nem változott után négy, nyolc vagy 26 hetes H. pylori fertőzés Annak ellenére, hogy az M1 makrofágok általában fokozzák az MHC-II és kostimulációs molekulák, nem tudtunk kimutatni fokozott MHC -II vagy CD86 gyomor makrofágok vagy CD103 + dendritikus akár nem immunizált vagy immunizált egerek a fertőzés után. Ezzel szemben, in vitro inkubáció H. pylori az akut gyulladásos válaszokat, makrofágok jellemzően polarizált az M1 és gyakoroljon erős anti-mikrobiális hatásokat. Például fertőzés Salmonella typhimurium katalógusa vagy Listeria monocytogenes katalógusa indukál M1 polarizációja makrofágok, amely szükséges az irányítást a fertőzés [39]. A gyulladás jellemzi eltolódást makrofág polarizáció M2, amely elősegíti a szövetek gyógyulását. Tuberkulózisos betegek megjelenítéséhez fokozott termelése Th2 citokinek, mint a fertőzés előrehaladtával [40], [41], amely képes indukálni M2 polarizációja makrofágok. Egereknél, M1 polarizációja makrofágok képezi korai válasz Mycobacterium tuberculosis katalógusa, míg alveoláris makrofágok polarizált M2 késő szakaszában fertőzés [42]. Azonban a váltás a makrofágok által indukált polarizáció krónikus fertőzések gyakran eredményez csökkentett kapacitás a makrofágok megölni a behatoló baktériumokat [39], [43]. Másrészt, a krónikus gyulladás tartós M1 makrofág polarizációt jár fokozott kockázata a rák kialakulásának. [4] katalógusa Van makrofágok hozzájárulnak, hogy a fogadó elleni védelem H. pylori katalógusa? Ellentétben a neutrofilek, makrofágok nem gyakran látható a gyomor lumenbe transzlokáció után az egész epithelium [44]. Mivel H. pylori Összefoglalva, ez a vizsgálat azt mutatja, hogy az oltás egerek ellen H. pylori katalógusa felerősíti M1 polarizációja gyomor makrofágok. Hasonló jelenség látható a humán atrófiás gasztritisz, ahol a vegyes M1 /M2 polarizáció jelen komplikációmentes gastritis helyébe egy M1-dominált polarizáció. Ez olyan daganatkeltő gyulladás, és a változó makrofág polarizáció M1 M2 ezért jelentenek terápiás célpont a krónikus H. pylori fertőzés katalógusa. Anyagok és módszerek katalógusa Etikai nyilatkozat katalógusa A vizsgálatot a kormány által jóváhagyott állat etikai bizottság (Göteborgs djurförsöksetiska nämnd, 328/2008 és 254 /2009). A regionális emberi etikai bizottság Västra Götaland megye Svédország (706/03 és 85/06) jóváhagyta a vizsgálatot, és írásos beleegyezésüket adták az összes résztvevő. Ez az egyetlen olyan hatóság, amely az etikai engedélyt embereken folytatott kutatás Svédországban, és ez közvetlenül nem kapcsolódó, a kórházak és az egyetemek. Katalógusa Egér katalógusa nőstény C57BL /6 egereket a Charles River Laboratories (Sulzfeld, Németország), vagy abban az esetben, a vakcinázási kísérletek Taconic (Ejby, Dánia). Az egereket korban 8-12 hét. Katalógusa A baktériumok és a fertőzés egerek katalógusa H. pylori egereknek négy 10 ul adag 500 ng H. pylori értékelése bakteriális kolonizáció mennyiségi értékelését a bakteriális kolonizáció immunizálási kísérletekben, az egyik fele a gyomrot óvatosan mossuk PBS-sel és homogenizáljuk Ultra Turrax homogenizálóban ( IKA Labortechnika, Staufen, Németország). Sorozathígításait a homogenizátum szélesztünk Skirrow agar lemezeken. Amikor gyomor lamina propria sejteket izoláltunk az egész gyomor mennyiségi becslésére gyomor bakteriális terhelés nem hajtható végre. Ebben az esetben, a gyomor, amely vágja a nagyobb görbület mentén, és PBS-ben mostuk, gyengéden szélesztettünk Skirrow agar lemezeken. A jelenléte H. pylori katalógusa telepek megerősítette egy ureáz teszt. Ily módon tudtuk megerősíteni H. pylori katalógusa kolonizáció és továbbra is használhatja az egész gyomor a sejtek elkülönítésre. katalógusa izolálása gyomor lamina propria sejtek katalógusa A mirigyes gyomrot vágva 5 mm-es darabokra, és inkubáltuk összesen háromszor H.
fertőzés áramlási citometriával és a valós idejű PCR-rel. Találtunk egy szekvenciális toborzása neutrofilek, eozinofilek és makrofágok a gyomor nyálkahártya fertőzött egerek. Génexpressziós analízise gyomor szövetek és rendezve makrofágok feltárta, hogy a gyomor makrofágok polarizált M1 után H. pylori
fertőzés, és ez a folyamat jelentős mértékben felgyorsult előzetes vakcinázás. Emberi H. pylori fertőzés katalógusa jellemezte vegyes M1 /M2 polarizációja makrofágok. Azonban a H. pylori katalógusa -asszociált atrófiás gastritis, a kifejezés indukálható nitrogén-oxid szintáz jelentősen emelkedett képest egyszerű gyomorhurut, azt jelzi, hogy a fokozott M1 makrofág polarizáció ebben pre-malignus elváltozás. katalógusa
felerősíti M1 polarizációja gyomor makrofágok, és hogy hasonló fokozott M1 polarizáció jelen van az emberi H. pylori katalógusa indukált sorvadásos gyomorhurut. katalógusa
megtelepedni a gyomor hámszövet több mint a fele a világ népességének [1]. A fertőzés gyakran élet-hosszú, és beindítja a krónikus gyulladás a gyomor nyálkahártya, ami körülbelül 1-2% a fertőzött egyének végül alakul gyomor adenokarcinóma [1]. Gyomorrák kialakulásával, különösen a bél típusú, egy többlépcsős folyamat, amely előrehaladtával évtizedek óta keresztül premalignus elváltozások a gyomor nyálkahártya, mint például a atrófiás gasztritisz, az intesztinális metaplázia, és diszplázia [2]. Az eredmény a fertőzés függ virulencia a fertőző H. pylori katalógusa törzs, a környezeti tényezők, mint a dohányzás és az étrend, és a fogadó genetikai tényezőket, amelyek befolyásolják a típusa és intenzitása a gyulladásos választ. [1] katalógusa
fertőzés és karcinogén kihívás, összehasonlítva a normál egerekben [6]. Míg iNOS hozzájárul a gyomorrák kialakulásával, a magas szintű a kemokin CCL18 gyomor tumorok társul túléléséhez gyomor rákos betegek [7]. Érdekes, iNOS által termelt klasszikusan aktivált /M1 makrofágok mivel CCL18 termelés fémjelzi a Alternatívaként aktivált /M2 makrofágok [8]. Összefoglalva, ezek az eredmények arra utalnak, hogy a makrofág polarizáció lehet fontos szerepet fejlődésének H. pylori
-asszociált gyomorrák.
fertőzés, makrofágok toborzott a gyomor nyálkahártya, amennyiben ezek hozzájárulnak a termelés a proinflammatorikus citokinek és kemokinek [10], [11], [12], [13], [14], [15] . Ezenkívül egy újabb vizsgálat azt mutatta, hogy a liposzóma-közvetített kiürülését makrofágok csökkent gyomor patológia H. pylori katalógusa-fertőzött egerekben [16]. Ennek ellenére, a funkció a makrofágok során in vivo H. pylori fertőzés katalógusa viszonylag gyengén meghatározott. A funkció makrofágok szorosan kapcsolt a polarizációs állapot, ami szintén úgy tűnik, hogy szerepe van a gyomorrák kialakulásával [6], [7]. Ezért megvizsgáltuk a makrofág polarizáció a gyomor nyálkahártyájában H. pylori
-fertőzött egerek és az emberek. Megmutatjuk, hogy az oltás egerek ellen H. pylori
sebesség és felerősíti az M1 polarizációja gyomor makrofágok. Ezen túlmenően, a rákot megelőző lézió atrófiás gasztritisz jellemzi fokozott makrofág M1 polarizáció emberekben.
Eredmények
fertőzés, mi fertőzött C57BL /6 egerekben az egér-adaptált H. pylori
Sydney törzs 1 (SS1), majd négy, nyolc és 26 hétig elemeztük a gyomor gyulladásos beszűrődés az egyes egerek többszínű áramlási citométerrel. A teljes száma lamina propria izolált sejtek a gyomorban nem változott az első négy hét a fertőzés, de nyolc héttel a fertőzés után az összes izolált sejtek megduplázódott, és 26 héttel a fertőzés volt egy nyolc-szeres növekedést a teljes száma izolált sejtek összehasonlítva a nem fertőzött egerek (1A.). Között a sejtek toboroznak a gyomor voltak makrofágok, eozinofil és neutrofil. Gyomor makrofágok azonosítottak expresszáló sejtek CD11b és a fő hisztokompatibilitási komplex II osztály (MHC-II), de hiányzik expressziója Gr1 (neutrophil marker), CD103 (által kifejezett egy részhalmazát dendritikus sejtek (DC-k)), és sziálsav-kötő immunoglobulin- mint lektin (Siglec-F, eosinophil marker) (ábra. 1B). Ezek a sejtek kifejezett makrofág marker F4 /80 (ábra. 1E), és az alapján sejt morfológiája is megerősítette a makrofágok (ábra. 1B). A frekvencia a makrofágok a gyomornyálkahártya változatlan maradt, miután négy és nyolc héttel a H. pylori fertőzés katalógusa (ábra. 1F). Azonban 26 hét után a gyakorisága gyomorrák makrofágok képest növekedett a nem fertőzött egerek (ábra. 1F). Katalógusa
dendritikus Továbbá, mivel ezek a sejtek szerepet játszanak, mint fontos antigénprezentáló sejtek nyálkahártyaszövetek [18]. Gyomor- CD103 + DC-k könnyen azonosítható festéssel CD11c és CD103 (ábra. 2B). A gyomor CD103 + DC-k magas szinten expresszált MHC-II, valamint állt a CD11b alacsony, és egy CD11b magas részhalmaza (ábra. 2C). Összehasonlításképpen, a gyomor CD103 - DC-ket minden CD11b magas (ábra. 2D). Ezen túlmenően, a CD103 + DC-k hiányzott expresszióját CD8α és F4 /80 (ábra. 2C). Azonban a gyakorisága CD103 + DS nem változott jelentősen a gyomornyálkahártya fertőzés után (ábra. 2E). Katalógusa
fertőzés a kifejezés a kostimulációs molekulák és az MHC-II által gyomor makrofágok és CD103 + DC-k, a ezen markerek expressziója elemeztük áramlási citometriával után négy, nyolc és 26 hétig a fertőzés. Az egyensúlyi állapotban, a makrofágok és a CD103 + DC-k a gyomor lamina propria kifejezte hasonló szinten a kostimulációs molekula CD86 valamint MHC-II (3A.). Meglepő módon, a kifejezés a CD86 és MHC-II akár gyomor- makrofágok vagy CD103 + DS-ben nem növekedett a fertőzés után H. pylori katalógusa relatív korú naiv egerek elemzett párhuzamosan (ábra. 3B). Így a makrofágok és a CD103 + DS a gyomor lamina propria nem upregulate CD86 és MHC-II után H. pylori fertőzés
ellenére folyamatos gyulladásos választ. katalógusa
fertőzés, használtuk real-time PCR mérésére gének expressziójának társított M1 vagy M2 polarizációja makrofágok a gyomor szövetben [8], [9]. Is mértük az IL-10, amely lehet előállítani szabályozó makrofágok [9]. Egyik markerek elemzett differenciáltan expresszálódó négy hét után H. pylori fertőzés
képest naiv egerek (4A.). Nyolc héttel a fertőzés után a kifejezés az M1-markerek iNOS és CXCL11 szignifikánsan emelkedett, és ezek a markereket tovább fokozódik a 26 hetes fertőzés (4A.). Ezen túlmenően, az IL-10-t serkentett nyolc és 26 hétig a fertőzés képest naiv egerek (ábra. 4A). Ezzel szemben, az M2 markerek találhatók a gyulladásos 1. zóna (FIZZ1) és argináz-1 nem eltérően expresszált a gyomorban négy, nyolc vagy 26 hetes fertőzés képest naiv egerek (ábra. 4A).
26 hétig (ábra. 4B). A mRNS expressziója iNOS és CXCL11 rendezett sejtpopulációk a fertőzött egerek, valamint a teljes gyomor lamina propria származó sejteket mind a naiv és a fertőzött egerek ezután határozzuk meg real-time PCR. Elegendő számú leválogatott makrofágok nem lehetett beszerezni naiv egerek megbízható elemzését mRNS expresszió. Gyomor makrofágok kifejezve a legmagasabb szintű iNOS és CXCL11 összehasonlítva a másik szortírozott sejtpopulációk és a teljes gyomor lamina propria sejtek válogatás előtt (ábra. 4C). Összefoglalva, ezek az eredmények azt mutatják, hogy a gyomorsav makrofágokat polarizált M1 alatt H. pylori fertőzés katalógusa.
. Ebből a célból, egereket immunizáltunk szublingválisan a H. pylori
lizátum és a kolera toxin adjuváns, és ezt követően megtámadta H. pylori
SS1. Ez immunizálási rend vezetett a jelentős csökkenéséhez bakteriális terhelés a gyomorban négy héttel a fertőzés után (ábra. 5A). Ugyanakkor, a kifejezés az M1 és M2 markerek a gyomorban analizáltuk real-time PCR-rei. Az immunizált egerekben, csak a kifejezés a CXCL11 szignifikánsan emelkedett négy héttel a fertőzés után összehasonlítva a naiv egerek (ábra. 5B). Ezzel ellentétben, immunizált egerek egy nagy fokozza a mind az M1-markerek iNOS és CXCL11 mivel a kifejezés az M2 markerek FIZZ1 és argináz-1 nem változott, négy héttel a fertőzés után (ábra. 5B). A fokozott expressziója iNOS és CXCL11 immunizált /fertőzött egerek nem volt köszönhető, hogy a immunizálás egyedül, hiszen immunizált de nem-fertőzött egerek nem változott a kifejezés bármely elemzett M1 vagy M2 markerek képest teljesen kezeletlen egerek (az adatokat nem mutatjuk ).
lizátum és a kolera-toxin, a makrofágok felhalmozódnak a gyomornyálkahártya és gyorsan polarizált M1 a fertőzés után.
és sorvadásos gyomorhurut makrofág polarizáció az emberi gyomor nyálkahártyáját. Ebből a célból, a kifejezés a humán M1 (iNOS, CXCL11) és az M2 markerek (CCL17, CCL18, CD206) mértük származó biopsziák antrumból real-time PCR-rei. H. pylori
-fertőzött egyének komplikációmentes gyomorhurut szignifikánsan megemelkedett expressziója mRNS mindkét M1 (iNOS, 8-szoros; CXCL11, 20-szeres) és M2 markerek (CCL17, 30-szeres; CCL18, 70-szeres, CD206 , 2-szeres), mint a nem fertőzött önkéntesekben (ábra. 6a). Vannak azonban olyanok, sorvadásos gyomorhurut (4/6 volt intestinalis metaplasia mellett sorvadás) kifejezve még magasabb iNOS mRNS képest a komplikációmentes gyomorhurut (20-szeres), míg a CXCL11 és markerek M2 polarizációs hasonlóképpen kifejezve (ábra. 6A). Valóban, a kifejezés iNOS 180-szorosára emelkedett egyének sorvadásos gyomorhurut képest nem fertőzött kontroll (6A.), Jelezve, fokozott M1 polarizációja gyomor makrofágok betegeknél sorvadásos gyomorhurut. Katalógusa
-fertőzött egyének kimutatható szintű volt az iNOS fehérje a antrumban mivel a koncentráció a iNOS volt a kimutatási határ alatt az összes nem fertőzött egyének (6B.). A kifejezés az iNOS mRNS és iNOS fehérje szignifikáns korrelációt mutatott (r 2 = 0,88, P
< 0,01), jelezve, hogy az mRNS elemzés tükrözi fehérje expresszió, még akkor is, ha a fehérje szintje alacsony. A koncentráció az M2 marker CCL18 növelte a gyomor szövet H. pylori katalógusa-fertőzött egyének képest nem fertőzött kontroll (6B.). Ráadásul az összefonódás a CCL18 fehérje szignifikáns korrelációt mutatott a kifejezés CCL18 mRNS (R 2 = 0,754, P katalógusa < 0,01). Katalógusa
-fertőzött egyének. Továbbá, sorvadásos gyomorhurut van társítva egy erős amplifikációját iNOS expresszió a gyomor nyálkahártyájának, jelezve egy továbbfejlesztett M1 polarizációja makrofágokban.
Discussion
. Megmutatjuk, hogy H. pylori
-fertőzött egyének kifejezni mRNS gyomornyálkahártya indikatív vegyes M1 /M2 polarizáció a makrofágok, és ezt tovább megerősítette a fehérje szinten. Azonban a H. pylori
-indukált atrófiás gasztritisz volt jelentős emelkedést a kifejezés a iNOS képest egyszerű gyomorhurut. Sorvadásos gyomorhurut ruházza fokozott a kockázata a gyomorrák képest egyszerű H. pylori
-asszociált gastritis [20]. A fokozott expressziója iNOS in atrófiás gasztritisz hozzájárulhat gyomorrák fejlesztés útján reaktív nitrogén gyökök, amely elősegítheti a karcinogenezis indukciós a DNS-károsodás, zavar a DNS-javítás, poszt-transzlációs módosítása, fehérjék, és a p53-mutációk [4]. Valóban, iNOS-hiányos egerek csökkentett előfordulási gyomor adenokarcinóma után H. pylori
fertőzés és kihívás egy kémiai karcinogén, összehasonlítva a normál egerekben [6]. Továbbá, polimorfizmusok promoter régiójában iNOS, ami magasabb transzkripciós aktivitás, korrelálnak nagyobb gyakorisággal az intesztinális típusú gyomorrák japán nők [21].
fertőzés, SS1-fertőzött C57BL /6 egerekben mutatott génexpressziós profil a gyomornyálkahártya indikatív makrofág M1 polarizáció. A génexpressziós analízise az szortírozott makrofágok izolált gyomornyálkahártya fertőzött egerek megerősítette, hogy expressziója az M1 markerek iNOS és CXCL11 gazdagodott a rendezett makrofág populáció. Továbbá, az M1-es polarizációs gyomor makrofágok jelentősen felgyorsult előzetes oltást. Már négy héttel a fertőzés után, immunizált egerekből serkentett expresszióját iNOS és CXCL11 egy hasonló szinten, mint ami látható 26 hét után a fertőzött csak-egerekben. Tanulmányok iNOS-deficiens egerekben kimutatták, hogy az iNOS elősegíti fejlődését atrófia és a rák, a gyomor nyálkahártya alatt Helicobacter
fertőzés [6], [22]. Ráadásul clearance H. pylori
védőoltás után függetlenül történik iNOS [23]. Így iNOS hozzájárulni látszik, hogy a fogadó patológiai helyett védelem alatt fertőzés H. pylori
. Ezért, ha a fokozott termelését iNOS a gyomornyálkahártya megmarad oltás után, lehet, hogy egy nemkívánatos mellékhatása, amelyek fokozhatják a súlyos H. pylori
indukált gyulladás és a rosszindulatú folyamatok, hacsak steril immunitás érhető el. Ugyanakkor a relatív hozzájárulását iNOS fogadó patológia versus védelem alatt különböző szakaszaiban H. pylori
fertőzés szavatolja a további vizsgálatot.
[15]. Ezen túlmenően, a számát makrofágok ismét emelkedett 60 és 120 nappal a fertőzés után képest naiv egerekben [15]. Mi kiterjeszteni ezeket az eredményeket, azt mutatja, hogy a felvételi ilyen veleszületett sejt populációk fennmarad egészen 26 héttel a fertőzés után. Emellett leírjuk a felhalmozási eozinofileket a gyomor nyálkahártya, egy sejtpopuláció, amely messze felülmúlják a gyomor makrofágok és neutrofilek (ábra. 1). Sőt, a szerepe eozinofil H. pylori katalógusa indukált gyomorhurut kell vizsgálni. Eozinofilek és makrofágok Share expressziója számos markerek, beleértve a CD11b és F4 /80 (1., [25]). Ezért több marker elemezni kell egyszerre annak érdekében, hogy megkülönböztetni ezeket a veleszületett sejt populációk. Katalógusa
. Ezzel szemben a mi tanulmány egy friss tanulmány szerint Kao et al. nem találtunk CD103 + DS gyomornyálkahártya nem fertőzött egerekben [26]. Inkább a CD103 + DS alakult 24 óra után H. pylori fertőzés
. Mivel az elemzés időpontjait különböznek, hogy a vizsgálati és a kísérleteink, nehéz közvetlenül összehasonlítani az eredményeket. Katalógusa
humán monociták [27], a humán monocita eredetű dendritikus sejtek [28], [29], [30], [31], rágcsáló csontvelő eredetű dendritikus sejtek [32], [33], vagy a humán primer gyomor- DC [34], serkenti upreguláció kostimulációs molekulák és MHC-II. Ezért az a tény, hogy fokozzák az MHC-II és a CD86 gyomor makrofágok és CD103 + dendritikus krónikus H. pylori
fertőzés okozhatja a gyulladásos miliő, és nem a baktériumok által önmagában. Sőt, a kölcsönhatás DS és H. pylori
in vitro nem szükségszerűen tükrözi, hogy mi történik in vivo, ahol a helyi mikrokörnyezetében a helyszínen az antigén beszerzés és az antigén prezentáció, valamint az egyenáramú részhalmaz részt vesznek az antigén bemutatása H. pylori katalógusa, befolyással lesz a válasz. Például, a szabályozó T-sejtek, amelyek elterjedtek a H. pylori
-fertőzött gyomornyálkahártya [35], [36], megakadályozhatja a túlszabályzását kostimulációs molekulák és az MHC-II DCS [37]. Ezen túlmenően, az IL-10 képes megakadályozni a túlszabályzását kostimulációs molekulák és az MHC-II a makrofágokon és DCS [38].
preferenciálisan tartózkodnak a gyomor nyálkahártya réteg vagy csatolt gyomornyálkahártya sejtek, makrofágok nem jön közvetlen érintkezésbe egész baktériumok. Valóban, kimerülése makrofágok által droggal feltöltött liposzómákat nem volt hatása a H. pylori katalógusa kolonizáció [16], ami arra utal, hogy a makrofágok nem közvetlenül járulnak hozzá a fogadó elleni védelem H. pylori
. Ezzel szemben a makrofágok elősegítik a gyomor patológia. Például liposzóma-közvetített kiürülését makrofágok enyhíthető a gyomorhurut által kiváltott H. pylori fertőzés katalógusa [16]. Ezen túlmenően, a szelektív törlését I-kB-kináz β mieloid sejtekben, ami megakadályozza, NF-kB ezekben a sejtekben, gátolta a gyomor atrófia után H. felis katalógusa fertőzés [45]. Így makrofágok úgy tűnik, nem járulnak hozzá Helicobacter katalógusa távolság, hanem inkább elősegíti a gyomor patológia. Katalógusa
SS1 növesztettünk Columbia ISO agarlemezeken 2 napon át 37 ° C-on, ami után azokat átadta a Brucella táplevesben kiegészített 5% magzati borjúszérumot (FCS) és antibiotikumokat (vankomicin, 10 ug /ml; B-polimixin , 20 U /ml, trimetoprim, 5 ug /ml), és inkubáljuk rázás éjszakán át 37 ° C-on mikroaerofil körülmények között. A fertőzés előtt, a motilitás a baktériumok igazoltuk mikroszkópia. A koncentráció a baktériumok becsültük spektrofotometriásán. Az egerek 3 × 10 8 telepképző egység SS1 intragasztrikusan.
Nyelvalatti immunizálás
lizátum (előállítva törzsből Hel305 a korábban leírt [46]) kombinált 10 ug kolera toxint (List Biological Laboratories Inc., Madison, NJ) szublingválisan 1 hetes időközönként [47], [48]. Két héttel az utolsó immunizálás után, az egereket gyomron és 3 × 10 8 telepképző egység H. pylori
SS1.
 Alkoholmentes zsírmájbetegség:Amit tudnia kell
Alkoholmentes zsírmájbetegség:Amit tudnia kell
 Steve SCD Healing Journal:26. hét – A kókuszvíz az anyatermészet gatorádája!
Steve SCD Healing Journal:26. hét – A kókuszvíz az anyatermészet gatorádája!
 Miért fordul elő savas refluxos fejfájás, és mit tehet?
Miért fordul elő savas refluxos fejfájás, és mit tehet?
 Hogyan befolyásolja a folsav a magas vérnyomást?
Hogyan befolyásolja a folsav a magas vérnyomást?
 A testmozgás és a bélmikrobióma
A testmozgás és a bélmikrobióma
 A gyógyszer megelőzheti a hasnyálmirigy-gyulladást emésztési eljárás után
A gyógyszer megelőzheti a hasnyálmirigy-gyulladást emésztési eljárás után
 Mik a Candida tünetei a bélben? 6 Okok
A Candida túlszaporodása a bélben olyan tüneteket okozhat, mint a hasmenés, hasi fájdalom, nyálka váladékozás a székletben, láz és fáradtság Candida Ez a gomba egy olyan típusa, amely általában a bőr
Mik a Candida tünetei a bélben? 6 Okok
A Candida túlszaporodása a bélben olyan tüneteket okozhat, mint a hasmenés, hasi fájdalom, nyálka váladékozás a székletben, láz és fáradtság Candida Ez a gomba egy olyan típusa, amely általában a bőr
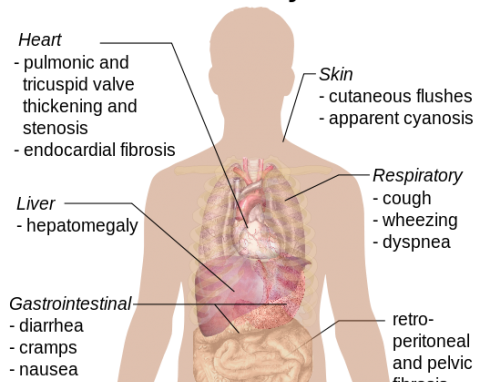 Karcinoid daganatok
A karcinoid daganat rosszindulatúnak minősül jól differenciált neuroendokrin sejtek (bioaktív vegyületeket termelő sejtek) proliferációja. A daganatok neuroendokrin szekréciós szemcséket tartalmazna
Karcinoid daganatok
A karcinoid daganat rosszindulatúnak minősül jól differenciált neuroendokrin sejtek (bioaktív vegyületeket termelő sejtek) proliferációja. A daganatok neuroendokrin szekréciós szemcséket tartalmazna
 Gastroenterológus a floridai Fort Pierce-ben
Ha egészségéről van szó, fontos, hogy olyan orvost válasszunk, aki nem csak képzett végzettséggel, hanem tapasztalattal és bizonyított múlttal rendelkezik a betegek sikeres kezelésében. Ebben a cikkbe
Gastroenterológus a floridai Fort Pierce-ben
Ha egészségéről van szó, fontos, hogy olyan orvost válasszunk, aki nem csak képzett végzettséggel, hanem tapasztalattal és bizonyított múlttal rendelkezik a betegek sikeres kezelésében. Ebben a cikkbe