Абстрактный
анализ содержимого желудков (SCA) и совсем недавно анализ стабильных изотопов (SIA) интегрирован с изотопным смешивания моделей стали обычным явлением методы исследования рациона и обеспечить понимание нагула экологии морских птиц. Тем не менее, оба метода имеют свои недостатки и отклонения, которые могут привести к трудностям в количественной оценке межгодовых и видоспецифические различия в рационе. Мы использовали эти два метода для одновременного количественного выращивания птенцов рацион антарктические ( Пигосселис Антарктида Введение Есть врожденные недостатки и уклоны при использовании SCA для количественной диеты морских птиц. Эта техника наиболее часто используется во время птенца выращивания, когда взрослые приносят еду на берег для своих птенцов; Таким образом, меньше известно о рационе морских птиц вне сезона размножения [2]. Содержимое желудка также отражает "снимок" недавней диете человека (8-16 часов) и может сильно различаться, что требует больших выборок статистически исследовать различия между видами, регионами и /или времени [3], [9], [ ,,,0],10]. Кроме того, SCA смещена в сторону последних диетических пунктов и добычи, которые не легко переваривается, таких как зоопланктон, и может недооценивать количество мягкотелых добычи, таких как рыба и кальмары [11], [12]. В то время как трудно добыча остается от содержимого желудка или гранул предоставить информацию о видовом составе добычи эти данные зачастую трудно интегрировать в общих оценках диеты состава [6], [8], [13]. Наши основные задачи заключаются в следующем:. 1) использовать одновременный сбор SCA и SIA сравнить возможности этих двух методов для выявления межгодовых и межвидовых различий в составе рациона в Пигосселис материалы и методы Этика заявление содержимое желудка, перо и добычи образцы Мы очищено перья, используя 2:1 хлороформ: метанол полоскание, высушенный на воздухе и нарезать их на мелкие осколки с ножницами из нержавеющей стали. Мы гомогенизируют целые образцы добычи, сушат их в течение 48 часов в печи при температуре 60 ° С и затем экстрагируют липиды из этих образцов с использованием аппарата Сокслета с 1:1 Петролеум-диэтиловый эфир: Этиловый эфир-диэтиловый эфир, смесь растворителей, в течение 8 часов [46]. Мы флэш-сгорает (Costech ECS4010 элементарный анализатор) примерно 0,5 мг каждого пера и добычи образца, загруженного в оловянных чашках и анализировали на содержание углерода и азота изотопов (δ 13C и δ 15N) через сопрягаемую Thermo Delta V Plus непрерывный поток масс-спектрометр соотношения стабильных изотопов (CFIRMS). Raw δ значения были нормализованы по двум-балльной шкале с использованием глутаминовой кислоты эталонных материалов с низкими и высокими значениями (т.е. USGS-40 (δ 13C = -26,4 ‰, δ 15N = -4,5 ‰) и USGS- 41 (δ 13С = 37,6 ‰, δ 15N = 47,6 ‰)). Образец точности на основе повторной выборки и эталонного материала составляла 0,1 ‰ и 0,2 ‰, для δ 13С, и б 15N соответственно. Статистический анализ содержание Желудок анализ два источника СИАР вариант модели, который используется P. antarcticum многоисточниковых модели SIAR Оба многоканальных исходные модели СИАР варианты (модели 3 и 4) предсказал, что антарктический криль составляли самую большую составляющую добычей антарктических и папуасских пингвинов куриных диет в 2008 году (таблица 4). Кроме того, обе модели нескольких источников Siar в целом согласились с двумя исходными СИАР модели оценки относительной доли криля против всех видов рыб, суммируется (таблицы 3 и 4). Тем не менее, наше первоначальное из нескольких источников СИАР модель (модель 3) была трудность оценки относительной доли отдельных видов рыб в обоих видов пингвинов куриных диетах в 2008 СИАР Model 95% интервалы Кредитоспособность 3 в целом перекрывается у разных видов рыб и средней относительной доли каждая из видов рыб отличались от оценок с использованием отолитовая водостойких массы (таблица 4, рис 3; антарктические:. χ 2 <суб> 5 = 62,65, р &л; 0,0001; Gentoo: χ 2 <суб> 4 = 41,70, р &л;. 0,0001) Обсуждение Желудок анализ содержания Два источника модели SIAR предсказывали относительно больший вклад рыбы выращивания птенцов диеты у обоих видов по сравнению с оценками биомассы SCA. Этот результат не является неожиданным, так как SCA, как полагают, недооценивают количество рыбы в рационе этих видов "в связи с пищеварением и Diel предубеждений, описанных выше [10], [12], [48]. Кроме того, модели двух источников SIAR также при условии, наименее переменных предсказания диеты композиции по сравнению с ГСС. SIA куриных перьев при условии, среднее значение рациона каждого отдельного цыпленка в течение всего времени роста перьев во время выращивания птенцов период [18], [19]. В отличие от этого, данные SCA представляют серию "SNAP-кадров" (в данном исследовании каждые 5 до 7 дней) пищи, что один из двух родителей кормить своего птенца [3]. Наше исследование показывает, что SIA тканей, которые интегрируют диеты в течение длительного периода времени являются врожденно менее изменчивы, чем СКА данного аналогичного размера выборки и являются более подходящими для изучения межгодовых различий в куриных диетах. Например, модели двух источников SIAR, используемые в нашем исследовании, были в состоянии идентифицировать межгодовую и видовым различия в относительной численности криля и рыбы в рационе не являются очевидными с помощью SCA. Когда предварительная информация на добычу видовой состав ограничен, например, вне сезона размножения, используя репрезентативную источник добычу в изотопных смешивания моделей может предоставить важную информацию о рационе морских птиц, когда мало что известно [24]. Тем не менее, наши результаты также показывают, что изменение видового состава добычи в пределах трофических или функциональных групп могут маскировать значительные различия в составе рациона, которые не будут очевидны из изотопных значений или смешивания предсказаний модели с использованием только представительные источников добычи. Этот результат был наиболее очевидным при изучении эффекта рыбьего добычи δ значения 15N на куриных перьев δ значений 15N и двух источников изотопный смешивания моделей, используемых в нашем исследовании. Например, антарктический пингвин куриных перьев δ значения 15N не отличались между годами (таблица 2). Кроме того, 95% интервалы доверия к диетической оценки из двух источников СИАР модели с помощью P. antarcticum Когда параметризуя наши две модели из нескольких источников Siar мы использовали отолитов данные для выбора соответствующих источников рыбы добычу включить в каждой видовой модели (Таблицы S1 и S2). Тем не менее, наше первоначальное из нескольких источников СИАР модель (модель 3) было трудно точно оценивающий индивидуальный видовой состав рыб части пингвинов диеты из-за общих черт в δ 13C и δ значения 15N среди многих видов рыб, включенных в качестве кормовой базы источников (рис. 3). видов антарктических рыб в целом потребляют криля и других видов рыб и из-за их подобного трофического уровня, эти виды рыб, как правило, имеют аналогичные δ значения 15N [45].
) и Gentoo ( P. Папуасов
) пингвины и методы изюминкой интеграции данных SCA повысить точность оценки диеты композиции с использованием SIA. Оценки биомассы SCA были весьма изменчивы и недооценили важность мягкотелых добычи, таких как рыба. Два источника, изотопные предсказания модели смешивания были менее изменчивы и определены межгодовую и видовым различия в относительных количеств рыбы и криля в рационе пингвинов не являются очевидными с помощью SCA. В противоположность этому, мульти-источник изотопных смешивания модели имели трудности, оценивающий диетический вклад видов рыб, занимающих аналогичные трофические уровни без уточнения с использованием SCA, полученных отолитов данных. В целом, наша способность отслеживать межгодовую и видовой специфические различия в пингвина диет с использованием SIA была расширена за счет интеграции данных SCA изотопным режимов смешивания тремя способами: 1) выбор подходящих источников добычи, 2) весовые комбинации изотопно подобной добычи в двух -source смешивания моделей и 3) переработка предсказывал вклад изотопно аналогичной добычи в моделях нескольких источников
<р> Цитирование:. Политыло MJ, Трайвелписа WZ, Карновского NJ, Ng E, Паттерсон WP, Emslie SD (2011) Интеграция Желудок Содержание и стабильный изотоп Анализы на Количественно диетах Pygoscelid пингвинов. PLoS ONE 6 (10): e26642. DOI: 10.1371 /journal.pone.0026642
<р> Редактор: Андре Chiaradia, Phillip Island Nature Parks, Австралия
<р> Поступило: 2 июня 2011 года; Принято: 30 сентября 2011 года; Опубликовано: 28 октября 2011
<р> Это статья с открытым доступом, свободной от всех авторских прав, и могут быть свободно воспроизведены, распространять, передавать, изменять, построен на, или иным образом использоваться кем-либо для любой законной цели. Работа доступна под общественное достояние Creative Commons cc0
<р> Финансирование:. Это исследование было профинансировано Программой США AMLR (http://swfsc.noaa.gov) и Национальный фонд Управления полярных программ США науки (www.nsf.gov) гранты С. Emslie и В. Трайвелписа. Доноры не играет никакой роли в дизайн исследования, сбора и анализа данных, решение о публикации или подготовки рукописи
<р> Конкурирующие интересы:.. Авторы заявили, что не существует никаких конкурирующих интересов
<р> анализ содержимого желудков (SCA) является одним из наиболее распространенных методов анализа диетического и дает представление о нагула экологии морских птиц и распределения, численности и демографии их добычи [1], [2]. Ранние исследования часто участвуют жертвоприношения животных для изучения содержимого желудка [3], в то время как в настоящее время неразрушающий, но до сих пор инвазивная, техника "промывание", чтобы заставить срыгивание обычно применяется [4], [5]. Когда Восстановленные содержимое желудка относительно непереваренной, можно оценить состав и частота встречаемости видов хищных и часто измеряют, взвешивают и секс индивидуальная добыча [3]. Кроме того, выявление и измерение трудно добычу остатки, такие как клювов кальмаров и отолитам, может предоставить информацию о размерах и массе хищных видов, когда добыча была частично или полностью переварили [6], [7], [8].
<Р> Последние достижения в области стабильного изотопа анализ (SIA) и изотопные смесительные модели показали большие перспективы в количественной оценке диетического состава морских птиц [14], [15]. Изотопные анализы основаны на концепции, что животные "то, что они едят" с тканевой стабильного азота (δ 15N) и углерода (δ 13С) коэффициентов, отражающих рацион питания во время синтеза [16]. Например, перья метаболически инертными после синтеза, так что перья из формирующихся возраста птенцов интегрировать диетическое историю во время выращивания птенцов период, как перья заменить натальной вниз [17], [18], [19]. Изотопные смешивания модели используют геометрические или байесовской процедуры для восстановления рациона питания животных на основе 13C и б δ 15N значения потребительских тканей и изотопно различных источников питания [20], [21]. SIA и изотопные смесительные модели имеют потенциал, чтобы обеспечить относительно неинвазивные и экономически эффективные количественные оценки диеты морских птиц на протяжении большей части их годового цикла [22], [23], [24].
<Р> Есть ограничения с использованием SIA для количественного определения диеты морских птиц. Когда изотопные подписи хищных видов, занимающих аналогичную трофического уровня перекрытия, например, в кормовой рыбы, перекрытия, это может быть трудно оценить их относительный вклад в потребительские диеты [25], [26]. Изотопные смешивания модели только полезны, так как данные, которые входят в них, требуя предварительного понимания возможных источников добычи и их отличительные изотопных значений [15]. Во многих случаях, предварительная информация отсутствует, и все возможные источники добычи не могут быть легко идентифицированы [24]. Когда все копытных значения изотопных не доступны, "представитель" виды часто используются или несколько источников объединены априори
для каждого трофического или функциональной группы [24], [27], [28]. Кроме того, в то время как исследования диет с использованием морских птиц SIA становятся обычным явлением, несколько исследований сравнивали одновременно количественные оценки состава диеты между СКА и SIA [28], [29]. Кроме того, это также общие для сравнения данных SIA на частоту SCA жертвы появления вместо более подходящих средств массовой основе оценок диеты композиции, полученных из СКА [30], [31], [32].
<Р> В это исследование мы одновременно количественно выращивания птенцов диета состав симпатрически разведения морских птиц, антарктического ( Пигосселис Антарктида
) и Gentoo пингвина ( P. папуасов
) в течение двух сезонов размножения на мысе Ширрефф, Ливингстон, Антарктида (62 ° 28'S, 60 ° 46'W), используя как SIA и SCA. Как и в других антарктических морских птиц, Пигосселис
пингвин диеты, как правило, состоит из зоопланктона, в основном антарктического криля ( Euphausia Суперба
), и мягкотелые, высших трофических хищных видов, таких как рыбы [ ,,,0],33]. Как выращивания птенцов диеты хорошо изучены с помощью присадок SCA на этом сайте, он обеспечивает прекрасное исследование для сравнения с SIA [34], [35], [36]. Мы стремимся, чтобы лучше понять относительные достоинства обоих методов и выделить использование SCA, чтобы сообщить изотопные модели смешивания для более точной количественной рацион морских птиц с использованием SIA
пингвина цыплят, 2) сравнить прогностическую способность двух источников (криль vs. рыба) линейная модель смешивания среди тех, с использованием репрезентативного видов рыб, и те, используя априорно
усредненные видов и год конкретных значений рыбы, и 3) оценить метод апостериори
интегрирующей СКА данные, чтобы лучше разъяснить Систематический состав рыбы части диет с использованием байесовской модели смешивания нескольких источников.
<р> использование животных в данном исследовании был проведен в соответствии с утвержденными протоколами использования животных из Калифорнийского университета в Сан-Диего Institutional уходу и использованию животных комитета (S05480) и в соответствии с Законом об Антарктике по сохранению разрешений обеспечивается Национальным научным фондом США С. Emslie (2006-001) и R. Холт (2008-008).
<р> Полевая состоялся в январе и феврале 2008 года и 2009 года в колонии примерно 4500 размножающихся пар антарктического пингвина и 800 гнездящихся пар пингвинов Gentoo на мысе Ширрефф. Мы собрали образцы желудка содержимого во время выращивания птенцов период после того, как птенцы достиг стадии CRECHE (&Гт 2,5-недельного возраста). Мы пробовали 2-5 уникальных племенных взрослых, возвращающихся из нагула поездок между 15: 00-17: 00 по местному времени в 5 до 7-дневными интервалами в общей сложности 10-14 Gentoo пингвины и 30 антарктические пингвины каждый год. Мы использовали технику водно-разгрузку после модификации Программы АНТКОМ по мониторингу экосистем (СЕМП) Стандартные методы [37]. В частности, мы не анализировали полное содержание желудка; а мы взяли примерно половину (около 350 г). Большая часть пищи под этой верхней части сильно переваривается и трудно объективно отделить от хищных видов и ее включение могут исказить как жертва идентификации и диеты оценки состава [10], [38]. Кроме того, мы оправдать этот метод выборки, как родители обычно не кормят всю свою нагрузку еды к птенцам [39], [40]. Избыток жидкости удаляли из каждого образца желудка, напрягая его через мелкое сито перед взвешиванием для получения массы образца (сырого веса). Из этих образцов, мы определили процент криля, рыбы и других материалов по частоте встречаемости и веса. Мы выздоровел рыбы отолитам из образцов диеты с помощью закрученного образцов в темном дном кастрюли и определили отолитам до минимально возможного таксономическом уровне с использованием внутреннего сбора справочной и опубликованную отолитов руководство [41]. Мы рассчитали возникновение частоты и минимальное количество особей (MNI) каждой рыбы таксонов в соответствии со стандартными методами [42]. Специально, мы оценили MNI путем суммирования большего числа вправо или влево отолитам с половиной количества эродированных отолитам неизвестной стороны, чтобы обеспечить консервативную оценку общего MNI, представленного в каждом образце желудка [42]. Кроме того, мы использовали отолитов измерения и опубликованные уравнения регрессии для расчета общей и процентов от общей массы водостойких для каждой рыбы таксона (таблица S1) [7], [13], [41], [43]. Из-за большого количества малых Pleuragramma antarcticum
отолиты, мы измерили случайную подвыборки 20-75 P. antarcticum
отолитам на пробу и использовали эти значения для оценки водостойких массы для этого вида.
<р> В феврале каждого года, мы собрали три грудные перья из случайной выборки 18-20 формирующихся цыплят каждого вида в то время как они готовились покинуть свои натальной колонии на море в 7-10 недель. С 2005 по 2009 год мы собрали репрезентативные образцы видов пингвинов во время добычи тралами, проведенных вдоль Южных Шетландских островов и северной части Антарктического полуострова и держали образцы замороженных перед анализом. Кроме того, мы дополнили эту библиотеку добычу с опубликованными изотопных значений двух рыб хищных, Protomyctophum bolini
и Champsocephalus gunnari
[44], [45].
Стабильный Изотопный анализ
<Р> Стабильные изотопные отношения выражаются в б обозначениях в единицах промилле (‰), в соответствии со следующим уравнением: где Х 13C или 15N и R является соответствующее отношение 13C / 12C или 15N / 14N. R
<суб> стандартные значения были основаны на Венской PeeDee белемнита (VPDB) при б 13С и атмосферная N <югу> 2 при б 15N.
изотопных смешивания моделей
<р> Мы использовали четыре модели варианта СИАР байесовской модели смешивания [21] в R среде (R Development Core Team 2007), чтобы изучить нашу способность количественно оценить рацион птенцов состав (Таблица S2). СИАР модельные оценки распределения вероятностей нескольких вкладов источника к смеси при учете наблюдаемой изменчивости в источнике и смеси изотопных подписей, диетического изотопного фракционирования и элементарной концентрации. Мы использовали две модели СИАР варианты с двумя источниками добычи (антарктического криля против "рыбы"), чтобы оценить состав диеты для каждого вида /комбинации год, используя δ 13C и δ 15N значения куриных перьев. Модель 1 использует б 13С и б 15N значений одного вида рыб, представитель P. antarcticum
, которая обычно находится в Пигосселис
пингвина диеты в качестве источника "рыба" [6]. Модель 2 использует вид и год удельная "рыба" δ 13C и δ значения 15N, вычисленные путем усреднения б 13C и б 15N значения нескольких видов рыб, взвешенных по их относительной процента водостойких массы рыбы (Таблицы S1 и S2).
<р> Мы использовали два дополнительных варианта смесительной модели СИАР с несколькими источниками добычи (6-7 в зависимости от видов пингвинов) для дальнейшей оценки методов интеграции данных содержимого желудка, чтобы лучше прояснить таксономического состав рыбы части пингвина диет. Для этих моделей, мы ограничили наш анализ цыплят данные пера с 2008 года, когда рыба часть куриных диет была самой разнообразной. Модель 3 является исходная модель из нескольких источников оценки относительного вклада криля ( E. ЗирегЬа
) и все виды рыб в нашей библиотеке добычи идентифицированы из отолитам в содержимого желудка каждого вида '(Таблицы S1 и S2). Модель 4 представляет собой апостериорной
информированного модель, в которой мы ограничили в результате задняя притягивает к тем, в которых относительная значимость отдельных видов рыб оценивался по обилию каждого вида, выявленных в ходе анализа отолитового. Для модели 4, мы ограничились задним черпает только те, где оцененные пропорциональные вклады наиболее распространенных рыб хищных на основе водостойких массы рыбы было больше, чем предполагаемые пропорциональных взносов второго наиболее распространенных рыб хищных, так и для второй наиболее распространенных рыб хищных больше, чем третьим наиболее распространенным и так далее для всех видов рыб. Для обеих начальных (модель 3) и информированных моделей (Model 4) из различных источников, мы также подвели итоги по всей рыбы и добычи оценивается пропорциональный вклад каждого вида рыб в части рыбы (т.е. исключая криль) пингвинов диеты. Для всех моделей Siar мы включили Пигосселис
пингвин перо δ 15N и δ факторы дискриминации 13С [47] и побежал 1 миллион итераций, поредели на 15, с начальным отбрасывания первого 40000. в результате в 64,000 задней рисует.
<р> Статистические расчеты были проведены с использованием SAS (версия 9.1). Мы проанализировали данные SCA для проверки различий между годами и видов, используя отдельные обобщенные линейные модели (Proc Genmod). Мы использовали биномиальное распределение ошибок и функцию логит связи для обобщенных линейных моделей с процентным соотношением (по влажной массе) или появление частоты каждого из наших трех основных добычи группы (криля, рыбы, а также «другой» добычу) в качестве переменных отклика. Для моделей, которые использовали MNI рыбы и реконструированных рыбы массы на пробу в качестве переменных отклика, мы использовали распределение Пуассона-ошибки с функцией логит связи. Для всех обобщенных линейных моделей, которые мы провели ретроспективном анализе с использованием коррекции Бонферрони и сообщил хи-квадрат и р-значения из тестовых статистик отношения правдоподобия для 3-го типа тестов.
<Р> Чтобы проверить различия в куриных перьев б 13C и δ значения 15N мы использовали многомерный анализ отклонений (MANOVA) наряду со значениями множественных сравнений Тьюки-Крамера разных видов и лет с использованием PROC ANOVA. Мы использовали аналогичную MANOVA для изучения б 13C и δ 15N значения видов в нашей библиотеке добычи. Мы использовали модели 95% интервалов доверия к сравнить оценки криля против рыбы среди двух исходных вариантов СИАР модели (модели 1 и 2) и ЗОБ влажной массы, а также процент вклада отдельных видов рыб для рыб части куриных диет среди мульти- варианты источника СИАР модели (модели 3 и 4) и SCA отолитовая полученных водостойких рыбы массы. Для облегчения прямого сравнения между моделями Siar и АСС, мы рассчитали средние Байеса и 95% интервалов доверия к каждому набору данных SCA с использованием цепи Маркова моделирования методом Монте-Карло (MCMC) через WinBUGS (версия 1.4). Эти моделирования MCMC были реализованы с использованием неинформативных Дирихле до с одинаковым числом итераций, редеет, и выкидывает, как наш СИАР модели анализа. Кроме того, мы использовали хи-квадрат СОГЛАСИИ тестов, чтобы сравнить распределение средних оценок процента вклада отдельных хищных видов рыб в рационе среди нескольких источников моделей SIAR вариантов и данных SCA.
<Р> Данные были исследованы на нормальности и равной дисперсии, все тесты были двусторонними и значение принималось на уровне 0,05. Стабильные значения изотопные куриных перьев и хищных видов представлены ± стандартное отклонение (SD), в то время как оценки диеты композиция из анализа содержимого желудка представлены ± стандартная ошибка (SE) в таблицах и ± 95% интервалы доверия к цифрах.
Результаты
<р> антарктический пингвин образцы желудка имели более высокий процент вклада относительно криля в образцах Gentoo пингвина (таблица 1а; χ 2 <к югу> 1 = 10,91, р = 0,0010). Тем не менее, мы не обнаружили никаких различий по годам (χ 2 <к югу> 1 = 0,22, р = 0,6375) или вида * год взаимодействия (χ 2 <к югу> 1 = 0,00, р = 0,9805). Кроме того, Gentoo образцы пингвина содержали значительно более высокий процент вклада рыбы по отношению к Чинстрап образцов пингвина, (χ 2 <к югу> 1 = 12,24, р = 0,0005), но мы не смогли обнаружить различия по годам (χ 2 <суб> 1 = 0,08, р = 0,7755) или вида * год взаимодействия (χ 2 <югу> 1 = 0,26, р = 0,6078). Процент вклад в образцах желудка других хищных видов, в том числе головоногих, амфипод и других видов эвфаузиевых не отличаются друг от вида пингвина (χ 2 <к югу> 1 = 0,00, р = 0,9694), год (χ 2 <югу> 1 = 0,36, р = 0,5468) или вида * год взаимодействия (χ 2 <к югу> 1 = 0,00, р = 0,9694).
<р> Мы нашли доказательства криля во всех антарктический пингвин образцы и во всех, кроме одного пингвина образца Gentoo (таблица 1а). Мы нашли доказательства рыбы во всех пробах пингвинов Gentoo и в 36.7-50.0% от Чинстрап образцов пингвина, даже если не было обнаружено влажной массы рыбы (табл 1а). Тем не менее, частота встречаемости рыб в Чинстрап образцах пингвина не отличались по годам (χ 2 <к югу> 1 = 1,09, р = 0,2966). Точно так же, частота встречаемости других видов хищных не отличались пингвин видов (х 2 <суб> 1 = 1,57, p = 0.2107), год (χ 2 <к югу> 1 = 0,00, р = 0,9481 ), или вид * год взаимодействия (χ 2 <к югу> 1 = 0,00, р = 0,9481).
<р> MNI рыбы и восстанавливали рыбы масса каждого образца отличались между видами и лет (Таблица 1b ). Антарктический пингвин диеты имели более низкий MNI и водостойких массу рыбы, чем пингвины Gentoo (MNI: χ 2 <к югу> 1 = 959,14, р &Лт; 0,0001; водостойких масса: χ 2 <к югу> 1 = 959,14, р &лт; 0,0001 ). Через видов и лет Антарктические пингвины имели более высокие водостойких рыбы массы в образцах диеты в 2008 году по сравнению с 2009 годом, в то время как Gentoo пингвины были и более низкую MNI и реконструированных рыбы массы в 2006 г. по отношению к 2009 году (MNI: χ 2 <к югу> 1 = 147,74 , р &л; 0,0001; восстанавливали масса: χ 2 <к югу> 1 = 1122,46, р &л; 0,0001). В общей сложности 96,3% всех отолитам были идентифицированы по крайней мере, на уровне рода, с шестью и пятью таксонов рыб, представленных в Чинстрап и Gentoo пингвина диет образцов, соответственно (таблица S1).
изотопные подписи куриных перьев и добыча
<р> Мы нашли δ 15N и δ 13C значения пингвина куриных перьев различались по видам (Уилкса λ, р ≪ 0,0001), год (Уилкса Х, р = 0,0409) и имел значительная вид * год взаимодействия (λ Уилкса, р &л; 0,0001). Gentoo пингвина цыплят была выше перо δ значения 15N чем антарктический пингвин цыплят в оба года (таблица 2, рис. 1). Тем не менее, в то время как Gentoo пингвина куриных перьев δ значения 15N были выше в 2009 г. по сравнению с 2008 годом, антарктический пингвин куриных перьев δ значения 15N не отличались между годами. Антарктические и Gentoo пингвина птенцы имели аналогичное перо δ 13C значения в 2008 году, но ниже и более высокие значения для антарктических и папуасских пингвинов в 2009 году, соответственно (таблица 2). Мы нашли δ 15N и δ 13C значения видов в нашей библиотеке пунктов добычи общих пингвина также значительно различались (λ Уилкса, р &Лт; 0.0001). Δ 15N и δ значения 13C сильно отличались от криля и рыб, в то время как значения изотопных перекрываются среди многих видов рыб (таблица 2, рис. 1).
Два источника Siar моделей
изотопные значения в качестве представителя "рыба" источник (модель 1) и вариант, который использовал в год и по конкретным видам, взвешенное "рыба" изотопные значения (модель 2) как предсказывали, что Gentoo пингвина птенцов потребляются относительно меньше криля и многое другое рыбы, чем антарктические пингвины в оба года (таблица 3). Тем не менее, при рассмотрении модели 95% интервалов доверие этих двух вариантов модели различаются по их способности определять видовой специфичностью, межгодовые различия в составе рациона. В то время как два источника варианта СИАР модели предсказывали, что Gentoo пингвина куриных диеты содержали высокий процент криля в 2008 году только 2 модели обнаружено большее количество рыбы в антарктический пингвин куриных диет в течение 2008 года по сравнению с 2009 г. (таблица 3). Два источника варианты модели SIAR предсказывали более высокий вклад рыбы в куриных диетах обоих видов пингвинов по сравнению с оценками диеты состава, полученных из SCA влажной массы (рис. 2). Оценки SCA также были более изменчивы, чем модели предсказания Siar для Gentoo пингвина куриных диет. Кроме того, SCA, полученные оценки среднего вклада криля и рыбы в рационе обоих видов "упали за пределами наших моделей двух источников Siar 95% верхние и низкие интервалы достоверности, соответственно (рис 2,. Таблицы 1 и 3).
<р> В отличие от этого, апостериорной
проинформировал нескольких источников модель СИАР (модель 4) лучше, чем у исходной модели СИАР нескольких источников (модель 3) на оценку видовой состав рыб части куриных диет. В то время как предсказание Модель 4 по средней относительной доли каждого вида рыб в антарктический пингвин куриных диетах незначительно отличались от оценок от отолитового разведенной массы (х 2 <суб> 5 = 14,55, р = 0,0125), полученные в результате интервалы Кредитоспособность 95% были снижены на 53,5 ± 17,2% по сравнению с Model 3 (диапазон: 33.2-82.2%, таблица 4, рис. 3). Кроме того, модель 4-х прогнозирования среднего удельного веса каждого вида рыб в рационе пингвинов папуа цыплят была аналогична оценкам отолитовая водостойких массы (χ 2 <суб> 4 = 3,40, р = 0,4949). Кроме того, полученные интервалы Кредитоспособность 95% были снижены на 52,0 ± 27,7% по сравнению с Model 3. (Диапазон:. 17.4-76.2%, таблица 4, рис 3)
<р> Наш анализ SCA выдвигает на первый план некоторые из возможных предубеждений, присущих при использовании этого метода. По аналогии с предыдущими исследованиями на мысе Ширрефф, мы наблюдали доказательства рыб, таких как отолитам, весы и линзы во многих образцах антарктического пингвина даже тогда, когда не было никакого измеримого количества ткани рыб влажной массы [34], [36]. Эти данные показывают, что биомасса рыбы потребляется взрослыми переваривает полностью до их возвращения в гнездовой колонии или, более вероятно, будет доставлен птенцов в сильно переваренной компонента содержания взрослого желудка, которые не могут быть объективно количественно [6], [10] , Кроме того, поскольку мы собрали образцы желудка во второй половине дня, наша выборка не включает в себя взрослых людей, которые промышляют в ночное время и, как правило, имеют гораздо более высокий процент и появление рыб в их образцах желудка [34], [48]. Кроме того, оценки диеты композиции, полученные из ЗОБ в нашем исследовании, были часто сильно варьирует, что делает его трудно обнаружить различия между годами и видов пингвинов (таблица 1). Этот вывод не представляется быть уникальным в морских птиц пищевых исследований с использованием SCA, которая часто требует высокого размера выборки и больших различий между группами в целях выявления межгодовой или видовым различия в составе рациона [3], [9]. Тем не менее, наше исследование показывает, что анализ отолитов еще может предоставить подробную информацию о видах специфической и временной изменчивости потребления хищных видов рыб, когда общие оценки Состав пищи происходит от содержимого желудка влажную массу менее информативны.
Два источника, модели SIAR
в качестве источника рыбы добычу (модель 1) перекрываются между годами (табл 3). В отличие от 95% интервалов доверия к двух источников модели СИАР с использованием ежегодно и взвешенные значения "рыба" конкретных видов (модель 2) предполагают большее обилие рыбы в антарктический пингвин куриных диет в 2008 году по сравнению с 2009 годом, что было подтверждено отолитом получена, средняя масса рыб Водостойких. В 2008 году рыба часть антарктического пингвина куриных диет состоит из шести видов рыб с ориентировочной б значения 15N 7,9 ± 0,7 ‰, а P. antarcticum
(δ 15N: 9,4 ± 0,5 ‰) был единственный вид рыбы в 2009 году диеты (табл S1 и S2). Хотя это 1,5 ‰ разница мала по сравнению с 4.6-6.1 ‰ различия между рыбой и крилем, этого было достаточно, чтобы посрамить межгодовые сравнения антарктический пингвин куриных диет в нашем исследовании.
из нескольких источников, модели SIAR
 Расстройства пищеварения:удивительные причины, почему у вас газы
Сколько газа слишком много? Газ в норме. Это часть вашего пищеварения, и она есть у всех. Большинство людей выделяют газы от пяти до 15 раз в день. Но если вы чувствуете, что у вас больше газа, чем
Расстройства пищеварения:удивительные причины, почему у вас газы
Сколько газа слишком много? Газ в норме. Это часть вашего пищеварения, и она есть у всех. Большинство людей выделяют газы от пяти до 15 раз в день. Но если вы чувствуете, что у вас больше газа, чем
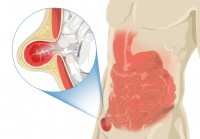 Паховая грыжа
На этой странице: Что такое паховая грыжа? Насколько распространены паховые грыжи? У кого чаще возникает паховая грыжа? Каковы осложнения паховых грыж? Каковы симптомы паховой грыжи? Что вызывает па
Паховая грыжа
На этой странице: Что такое паховая грыжа? Насколько распространены паховые грыжи? У кого чаще возникает паховая грыжа? Каковы осложнения паховых грыж? Каковы симптомы паховой грыжи? Что вызывает па
 Гастроэнтерология Сан-Франциско представляет наш портал для пациентов
Гастроэнтерология Сан-Франциско теперь предлагает портал для пациентов, который обеспечит нашим пациентам легкий доступ к нашему офису и услугам. После заполнения краткой регистрационной формы пациент
Гастроэнтерология Сан-Франциско представляет наш портал для пациентов
Гастроэнтерология Сан-Франциско теперь предлагает портал для пациентов, который обеспечит нашим пациентам легкий доступ к нашему офису и услугам. После заполнения краткой регистрационной формы пациент