obsah žalúdka analýza (SCA) a najnovšie stabilný izotop analýzy (SIA) integrovaný s izotopovými miešacích modely sa stali bežné metódy pre diétne štúdia a poskytnúť vhľad do hľadania potravy ekológie morských vtákov. Avšak, obe metódy majú nedostatky a predsudky, ktoré by mohli viesť k ťažkostiam pri kvantifikácii medziročný a druhovo špecifické rozdiely v diétach. Použili sme tieto dve metódy súčasne kvantifikovať chick odchovu stravu remienok ( Pygoscelis antarctica Citácia :. Polito MJ, Trivelpiece WZ, Karnovsky NJ, Ng E, Patterson WP, Emslie SD (2011) Integrácia Žalúdok obsah a stabilný izotop analýzy vyčísliť stravy Pygoscelid Penguins. PLoS ONE 6 (10): e26642. doi: 10,1371 /journal.pone.0026642 Editor: André Chiaradia, Phillip Island Nature Parks, Austrália prijatá: 02.6.2011; Prijaté: 30.Septembra 2011; Uverejnené: 28.října 2011 Toto je článok o otvorený prístup, bez všetky autorské práva, a môže byť voľne reprodukovať, distribuovať, prenášať, úprave, postavený na, alebo inak používať ktokoľvek, pre akýkoľvek zákonný účel. Práca je sprístupnený v rámci public domain venovanie Creative Commons CC0 Financovanie :. Tento výskum bol financovaný americkým AMLR programu (http://swfsc.noaa.gov) a US National Science Foundation Úradu Polar Programy (www.nsf.gov) granty S. Emslie a W. Trivelpiece. Platcovia mal žiadnu úlohu v dizajne štúdie, zber a analýzu dát, rozhodnutie publikovať, alebo prípravu rukopisu Konkurenčné záujmy: .. Autori vyhlásili, že žiadne konkurenčné záujmy neexistujú Úvod obsahová analýza žalúdka (SCA) je jednou z najčastejších metód pre diétne analýzu a poskytuje vhľad do hľadania potravy ekológie morských vtákov a rozdelenia, množstva a demografia ich koristi [1], [2]. Skoršie štúdie často zapojené obetovanie zvierat skúmať obsah žalúdka [3], zatiaľ čo v súčasnej dobe nedeštruktívne, ale napriek tomu invazívne, "výplach" technika vynútiť regurgitácia sa bežne používa [4], [5]. Pri obnovenej obsah žalúdka sú pomerne nestrávené, je možné odhadnúť zloženie a početnosť výskytu druhov koristi a často meria, váži a sex individuálne korisť [3]. Okrem toho, identifikovať a merať tvrdý korisť zvyšky, ako sú chobotnice zobáky a otoliths, môže poskytnúť informácie o veľkosti a hmotnosti druhov koristi, keď korisť bola čiastočne alebo úplne rozštiepi [6], [7], [8]. k dispozícii sú vlastné nevýhody a predsudky pri použití SCA kvantifikovať Seabird diéty. Táto technika bola najčastejšie používaný počas odchovu mláďaťa, kedy dospelí priniesť jedlo na breh za ich mláďatá; teda menej je známe o diéty morských vtákov mimo obdobia rozmnožovania [2]. obsah žalúdka tiež odrážajú "snímku" nedávne stravy jedinca (8-16 hodín) a môže byť vysoko variabilné, čo vyžaduje veľké veľkosti vzorky štatisticky posúdiť rozdiely medzi druhmi, regióny a /alebo času [3], [9], [ ,,,0],10]. Okrem toho SCA je tlačená smerom nedávnych potravinových položiek a korisť, ktorá nie je ľahko stráviteľné, ako zooplanktónu, a môže podceňovať množstvo mäkkým telom koristi, ako sú ryby a kalmáre [11] [12]. Kým tvrdé korisť zvyšky zo žalúdočného obsahu alebo pelety poskytovať informácie o dravých druhového zloženia sú tieto údaje často ťažké integrovať do celkovej odhady zloženie stravy [6], [8], [13]. Nedávne pokroky v stabilnom izotopom analýza (SIA) a izotopové zmiešavacie modely ukázali ako veľmi sľubný vyčíslenie zloženie potravy morských vtákov [14], [15]. Izotopové analýzy sú založené na predstave, že zvieratá ", je to, čo jedia" s tkanivom stabilný dusíka (δ 15 N) a uhlíka (δ 13C), ktoré odrážajú pomery stravy v dobe syntézy [16]. Napríklad, perie sú metabolicky inertné po syntéze, takže peria z začínajúcim veku kurčiat integrovať stravovacie návyky počas obdobia chick-správať ako perie nahrádzať pôrodníckej nadol [17], [18], [19]. Izotopové zmiešavacie modely používajú geometrické alebo bayesovských postupy rekonštruovať zvieracie stravy založenej na delta 13C a δ 15N hodnoty spotrebnej tkanív a deutériom odlišné potravinové zdroje [20], [21]. SIA a izotopové miešacej modely majú potenciál poskytnúť pomerne neinvazívna a nákladovo efektívne kvantitatívne odhady morských vtákov diét počas veľa z ich ročného životného cyklu [22], [23], [24]. Existujú obmedzenia použitím SIA kvantifikovať Seabird diéty. Keď izotopové podpisy dravých druhov, ktoré zaujímajú podobný trofické úrovne prekrytia, napríklad v kŕmnej ryby, prekrývajú, môže byť ťažké odhadnúť ich relatívny podiel na spotrebiteľských diétach [25], [26]. Izotopové zmiešavacie modely sú len tak užitočné ako dáta, ktoré idú do nich, ktoré vyžadujú predchádzajúce znalosti o možných zdrojoch koristi a ich charakteristickými izotopové hodnoty [15]. V mnohých prípadoch, priori informácie chýba, a všetky možné zdroje korisť nemôže byť ľahko identifikovaný [24]. Keď sú všetky korisť hodnoty izotopov nie sú k dispozícii, "reprezentatívne" druhy sú často používané alebo viacerých zdrojov sú kombinované a priori v táto štúdia nám súčasne kvantifikovať chick odchovu stravy zloženie sympatrically chov morských vtákov sa Chinstrap ( Pygoscelis antarctica Naše hlavné ciele sú: a. 1) používať súčasný zber SCA a SIA porovnať schopnosť týchto dvoch metód pre detekciu medziročné a inter-špecifické rozdiely v zložení stravy v Pygoscelis materiály a metódy Ethics vyhlásenie využívanie zvierat v tejto štúdii bola vykonaná v súlade so schválenými protokolmi používajú zviera z University of California v San Diegu ústavnej zvierat starostlivosť a používanie výboru (S05480) a v súlade s antarktickej podľa zákona o ochrane povolenia stanovené americkou National Science Foundation S. Emslie (2006-001) a R. Holt (2008-008). Žalúdočné obsah, perie a korisť vzorky Práca v teréne sa uskutočnilo v januári a februári 2008 a 2009 v kolónii asi 4500 chovných párov remienok tučniakov a 800 chovné páry tučniakov osloch na myse Shirreff. Zhromaždili sme vzorky žalúdočného obsahu na dobu chick odchovu mláďat po dosiahla štádia jaslí (viac ako 2,5 týždňov veku). odber vzoriek sme 2-5 unikátny chov dospelým vracajúcim sa z zbierajúci ciest medzi 15: 00-17: 00 miestneho času v 5-7 denných intervaloch pre celkom 10-14 tučniak oslí a 30 remienok tučniaky každý rok. Použili sme vodu vykladanie techniky po zmene Monitorovacieho CCAMLR ekosystému Program (CEMP) Štandardné metódy [37]. Konkrétne sme nemali analyzovať celý obsah žalúdka; skôr sme sa približne jedna polovica (asi 350 g). Väčšina potravín pod touto hornej časti je ťažko stráviteľné a je ťažké objektívne oddeliť druhov koristi a jej začlenenie môžu ovplyvniť obe koristi identifikácie a diétne odhady zloženie [10], [38]. Ďalej ospravedlniť túto metódu odberu vzoriek ako rodičia zvyčajne nepodávajú celý svoj náklad potravín, ktoré pomáhajú kurčiat [39], [40]. Prebytočná kvapalina sa odstráni z každej vzorky žalúdka namáhanie skrz jemné sito pred vážením sa získa hmotnosť vzorky (hmotnosť za vlhka). Z týchto vzoriek sme stanovili percento krilu, ryby a iný materiál podľa výskytu frekvencií a hmotnosti. zotavil sme rybie otoliths zo vzoriek diét vírením vzorky tmavé dňom panvice a zistené otoliths na najnižšiu možnú taxonomickú úroveň pomocou vnútorného referenčného zber a publikované otolitových príručku [41]. Vypočítali sme výskyt frekvencie a minimálny počet jedincov (MNI) každého rýb taxónov nasledujúce štandardnými metódami [42]. Špeciálne sme sa odhaduje MNI ako súčet väčší počet buď vpravo alebo vľavo otoliths s polovičným počtom erodovaných otoliths neznámeho strane, aby konzervatívny odhad celkového MNI je v každom žalúdku vzorky [42]. Okrem toho sme použili otolitové merania a zverejnených regresnej rovnice pre výpočet celkom a percento celkovej hmoty riedené na každú zásob rýb taxónov dopravy (pozri tabuľku S1) [7], [13], [41], [43]. Vzhľadom na vysoký počet malých Pleuragramma antarcticum V februári každého roka, sme zhromaždili tri prsníka peria z náhodnej vzorky 18-20 začínajúcim mláďat všetkých druhov zatiaľ čo že sa pripravujú k odchodu ich rodných kolónie na mori na 7-10 týždňov veku. Od roku 2005 do roku 2009, sme zhromaždili reprezentatívne vzorky tučniaka druhov koristi v priebehu vlečných sietí vedených pozdĺž Južnej Shetlandy a severnej Antarktického polostrova a uchovávané vzorky zmrazené pred analýzou. Ďalej doplnené túto korisť knižnicu s publikovanými izotopov hodnotami dve ryby koristi, Protomyctophum Bolin stroje a Champsocephalus gunnari vyčistiť perie sme pomocou 2: 1 chloroform: metanol plákania, suší na vzduchu a nakrájajte na malé fragmenty s nožnicami z nerezovej ocele. homogenizuje sme celé vzorky korisť, suší sa im po dobu 48 hodín v sušiarni pri teplote 60 ° C a potom sa extrahuje lipidy z týchto vzoriek za použitia Soxhletovom prístroji s 1: 1 petroléteru: etyl-éter zmesi rozpúšťadiel počas 8 hodín [46]. My blikať-spaľovaný (cost ECS4010 elementárneho analyzátora) približne 0,5 mg každé perie a koristi vzorky vloženého do cínových pohárov a analyzované na uhlíka a dusíka izotopov (δ 13C a δ 15N) pomocou rozhraním Thermo Delta V Plus nepretržitý prúd stabilný izotop pomer hmotnostný spektrometer (CFIRMS). Surové delta hodnoty boli normalizované na dva-bodovej stupnici pomocou glutámovej kyseliny referenčných materiálov s nízkymi a vysokými hodnotami (tj USGS-40 (δ 13C = -26,4 ‰, δ 15N = -4,5 ‰) a USGS- 41 (δ 13C = 37,6 ‰, δ 15N = 47,6 ‰)). Vzorka presnosť založený na opakovanom vzorky a referenčného materiálu bola 0,1 ‰ a 0,2 ‰, pre δ 13C a ó 15N, resp. Stabilné pomery izotopov sú vyjadrené v ó zápise v jednotkách promile (‰), podľa tejto rovnice: kde X je 13C alebo 15 N, a R je zodpovedajúci pomer 13C / 12C alebo 15 N / 14N. R Použili sme štyri modelové varianty SIAR Bayesian miešanie v oblasti výskumu životného prostredia (R Development jadro tímu 2007) modelu [21], aby preskúmala našu schopnosť kvantifikovať chick zloženie stravy (tabuľka S2). V Siar Model odhaduje pravdepodobnosť rozdelenie rôznych príspevkov zdrojových k zmesi pri účtovaní zistených rozdielov v zdroji a zmes izotopov podpisov, diétne izotopovou frakcionáciu a elementárne koncentrácie. Použili sme dva SIAR varianty modelu s dvoma zdrojmi koristi (antarktického krilu versus "ryby") pre odhad zloženie stravy pre každý druh /rok kombinácii s použitím δ 13c a δ 15N hodnoty kuracích peria. Model 1 používa delta 13c a ó 15N hodnotami reprezentatívne druhy rýb, p. antarcticum sme použili dve ďalšie varianty zmiešavacie modelu SIAR s niekoľkými zdrojov koristi (6-7 v závislosti na tučniaky druhov), ďalej zhodnotiť metódy integrácie dát žalúdka obsahu pre lepšie objasnenie taxonomické zloženie rybie časti Penguin stravy. U týchto modelov, sme obmedzené naše analýzy, mláďa dát peria z roku 2008, kedy sa ryby časť kuracích diét bol najviac rôznorodé. Model 3 je počiatočná multi-source model, odhad relatívny prínos krilu ( E. Superba Štatistická analýza Štatistické výpočty boli vykonané pomocou SAS (verzia 9.1). Analyzovali sme dáta, SCA na testovanie rozdielov medzi jednotlivými rokmi a rybieho druhu za použitia samostatných zovšeobecnené lineárne modely (Proc Genmod). Použili sme binomické rozdelenie chýb a funkcie logit link na zovšeobecnené lineárne modely s percentuálne zloženie (o vlhké hmoty), alebo frekvencia ich výskytu každej z našej tri hlavné korisťou skupiny (krill, ryba, a "iný" korisť) ako hodnoty reakcie. Pri modeloch, ktoré používajú MNI rýb a obnovených rybie hmotu na vzorke, ako je premenných odozvy, sme použili rozdelenie Poisson-chyba s funkciou logit link. Pri všetkých zovšeobecnených lineárnych modelov sme vykonali post-hoc analýzy za použitia Bonferroniho korekcie a hlásené chi-kvadrát a P-hodnoty zo pravdepodobnosť pomer testovacích štatistík pre 3 typových skúšok. Ak chcete otestovať rozdiely v chick peria ó 13C a δ hodnoty 15N sme použili viacrozmerné analýzy rozptylu (Manová) spolu s Tukom-Kramer hodnôt Multiple porovnaní naprieč druhmi a rokov s použitím PROC ANOVA. Použili sme podobný Manovi skúmať ó 13c a δ 15N hodnoty druhov v našej koristi knižnici. Použili sme model 95% dôveryhodnosť intervaly porovnať odhady krilu vs. rýb medzi dve-source SIAR modelové varianty (modely 1 a 2) a SCA mokrá hmotnosť a percentuálny podiel druhov na rybie porciu kuracieho diét medzi multi- zdroj SIAR modelové varianty (modely 3 a 4) a SCA otolitových odvodený z riedeného rybie hmotu. Aby sa uľahčilo priame porovnanie SIAR modely a SCA sme vypočítali bayesovských priemery a 95% intervaly dôveryhodnosť pre každý SCA dátovom súbore pomocou simulácie Markov Chain Monte Carlo (MCMC) cez WinBUGS (verzia 1.4). Tieto simulácie MCMC boli vykonané za použitia non-informatívne Dirichlet predchádzajúcej s rovnakým počtom iterácií, tenší a blokuje ako naše SIAR modelovej analýzy. Ďalej sme použili Chi-kvadrát dobrotu testov fit porovnať distribúciu priemerných odhadov percentuálny podiel jednotlivých dravých druhov rýb do diét medzi multi-source Siar modelov variantov a údajov SCA. Údaje boli vyšetrené na normality a rovnaké rozptyl, všetky testy boli dvojchvostý a význam sa predpokladalo na úrovni 0,05. Stabilné hodnoty izotopové zloženie kuracích perie a dravých druhov sú prezentované ± štandardná odchýlka (SD), zatiaľ čo odhady zloženie stravy z analýzy obsahu žalúdka sú prezentované ± štandardná odchýlka (SE), v tabuľkách a ± 95% vierohodnosti intervaly v číslach. žalúdočného obsahu analýza tučniak čiapočkatý žalúdočné vzorky mali vyššie percento krilu vzhľadom k vzorkách tučniak somárov (tabuľka 1a; χ 2 1 = 10,91, p = 0,0010). Sme však zistené žiadne rozdiely podľa roku (χ 2 1 = 0,22, p = 0,6375), alebo druh * rok interakcie (χ 2 1 = 0,00, p = 0,9805). Rovnako tak Gentoo vzorky tučniak obsahovali výrazne vyšší percentuálny podiel rýb v pomere k vzorkách tučniak čiapočkatý (χ 2 1 = 12,24, p = 0,0005), ale nemohli sme zistiť rozdiely medzi jednotlivými rokmi (χ 2 1 = 0,08, p = 0,7755), alebo druh * rok interakcie (χ 2 1 = 0,26, p = 0,6078). Percentuálny podiel vzoriek žalúdka iných druhov koristi, vrátane hlavonožcov, amphipods a iné druhy euphausiid nelíšila od tučniakov druhov (χ 2 1 = 0,00, p = 0,9694), rok (χ 2 1 = 0,36, p = 0,5468), alebo druhy * interakcie ročne (χ 2 1 = 0,00, p = 0,9694). Zistili sme dôkazy o krilu vo všetkých tučniak čiapočkatý vzorky a vo všetkých prípadoch okrem jedného tučniak oslí vzorky (tabuľka 1a). Našli sme dôkazy o rybách vo všetkých vzorkách Tučniak somárov a 36.7-50.0% vzoriek tučniak čiapočkatý, aj keď tam bola zistená žiadna vlhká hmotnosť rýb (tabuľka 1a). Avšak, frekvencia ich výskytu rýb vo vzorkách tučniak čiapočkatý sa nelíšil naprieč jednotlivými rokmi (χ 2 1 = 1,09, p = 0,2966). Rovnako tak frekvenciu ich výskytu iných druhov koristi nelíšila Penguin druhov (× 2 1 = 1,57, p = 0.2107), rok (χ 2 1 = 0,00, p = 0,9481 ), alebo druh * rok interakcie (χ 2 1 = 0,00, p = 0,9481). MNI rýb a rekonštituovaný ryby hmotnosť na vzorke líšili medzi druhmi a rokoch (Tabuľka 1b ). Tučniak čiapočkatý diéty mal nižšiu MNI a riedené rybie hmotnosť ako tučniak oslí (MNI: χ 2 1 = 959,14, p 0,0001; rekonštituovaný hmotnosť: χ 2 1 = 959,14, P < 0,0001 ). Naprieč druhy a roky Chinstrap tučniaky mali vyššie opätovného zriadenia ryby hmotu vo vzorkách diétnych v roku 2008 v porovnaní s rokom 2009, zatiaľ čo tučniak somárov mali obaja nižší MNI a obnovených rybích mas v roku 2006 v porovnaní s rokom 2009 (MNI: χ 2 1 = 147,74 , p < 0,0001; rekonštituuje hmotnosť: χ 2 1 = 1122,46, p < 0,0001). Celkom 96,3% všetkých otoliths boli identifikovateľné aspoň na úrovni rodu, so šiestimi a piatich rýb taxónov, znázornené na remienok a Tučniak somárov diéty vzoriek, v danom poradí (tabuľka S1). Zistili sme, δ 15N a δ 13C hodnoty tučniakov kuracích perie líšila podľa druhov (Wilks 'λ, p 0,0001), rok (Wilks' lambda, p = 0,0409) a mal významný podiel tejto skupiny druh * interakcie rok (Wilks 'λ, p < 0,0001). Tučniak somárov mláďatá mali vyššie perie δ hodnoty 15N než remienok tučniak kurčiat v oboch rokoch (Tabuľka 2, Obr. 1). Avšak, zatiaľ čo Gentoo penguin chick perie δ hodnoty 15N boli vyššie v roku 2009 v porovnaní s rokom 2008, Chinstrap tučniak perníkový perie δ hodnoty 15N nelíšili rokov. Chinstrap a Tučniak somárov mláďatá mala podobnú perie δ 13C hodnoty v roku 2008, ale nižšie a vyššie hodnoty pre remienok a tučniak oslí v roku 2009, v uvedenom poradí (tabuľka 2). Zistili sme, δ 15N a δ 13C hodnoty druhov v našej knižnici spoločnej tučniakov položky koristi tiež významne odlišovali (Wilks 'λ, P &0,0001). Δ 15N a δ hodnoty 13C značne líšili medzi planktón a druhy rýb, zatiaľ čo hodnoty izotopov prekrývali z mnohých druhov rýb (tabuľka 2, obr. 1). dvoch-source SIAR modelu variant, ktorá používa p. antarcticum multi-source Siar modely Oba multi-source SIAR modelových variantov (modely 3 a 4) predpovedala, že antarktického krilu zahŕňal najväčší korisť zložku remienok a tučniak oslí kuracích diét v roku 2008 (tabuľka 4). Okrem oboch multi-source modely Siar v zásade súhlasil s dva-source SIAR modelovej odhady relatívneho podielu krilu vs. všetky druhy rýb zhrnul (tabuľky 3 a 4). Avšak, naše počiatočné multi-source SIAR model (Model 3) len s ťažkosťami odhad relatívny podiel druhov na oboch tučniak druhy kuracích diét v roku 2008. SIAR Model 95% vierohodnosti intervaly 3 vo veľkej miere prekryté cez druhov rýb a stredné relatívnym podielom každý druh rýb sa líšila od odhadov s využitím otolitových rekonštituovaný hmotnosť (tabuľka 4, obr.3; Chinstrap :. χ 2 5 = 62,65, p < 0,0001; Gentoo: χ 2 4 = 41,70, p. < 0,0001) na rozdiel od toho a posteriori Diskusia žalúdok obsahová analýza Naša SCA analýza poukazuje na niektoré z možných predsudkov spojených pri použití tejto metódy. Podobne ako pri predchádzajúcich štúdií na myse Shirreff sme pozorovali dôkazy o ryby ako otoliths, váha a šošovky v mnohých vzorkách tučniak čiapočkatý, aj keď nebolo žiadne merateľné množstvo rýb tkaniva vlhkej hmoty [34], [36]. To je dôkazom, že rybí biomasy spotrebovanej dospelých trávi úplne pred ich návrate do chovnej kolónie alebo, čo je pravdepodobnejšie, sa dodáva do kurčiat v silne štiepeného zložky dospelých obsahu žalúdka, ktoré nemožno objektívne [6] kvantifikovať, [10] , Okrem toho, pretože sme zhromaždili žalúdočných vzoriek počas neskorého popoludnia, náš vzorka nezahŕňa dospelých, ktorí pásli v noci a majú tendenciu mať oveľa vyššie percento a výskyt rýb v ich žalúdočných vzoriek [34], [48]. Okrem toho, zloženie stravy odhady odvodené od SCA v našej štúdii boli často veľmi variabilný, takže je ťažké odhaliť rozdiely medzi rokmi a tučniak druhov (Tabuľka 1). Toto zistenie sa nezdá byť jedinečný v morských vtákov diétnych štúdiách s použitím SCA, ktorá často vyžaduje vysokú veľkosťou vzorky a veľké rozdiely medzi oboma skupinami pri odhaľovaní medziročný alebo druhovo špecifické rozdiely v zložení stravy [3], [9]. Avšak, naša štúdia ukazuje, že analýza otoliths môže stále poskytovať podrobné informácie o druhovo špecifické a časové rozdiely v konzumácii rýb koristi pri celkovej odhady zloženie stravy odvodený od žalúdka obsahu Vlhká hmota sú menej informatívne. Two-source Siar modelov predpovedá relatívne väčší podiel rýb na Chick-chovu stravy u oboch druhov v porovnaní s odhadmi SCA biomasy. Tento výsledok nie je neočakávaný ako SCA je myšlienka podceňovať množstvo rýb v diétach týchto druhov 'kvôli trávenie a diel predsudky opísaných vyššie [10], [12], [48]. Okrem toho dva-source Siar modely tiež za predpokladu, že najmenej variabilné predpovede zloženie stravy v porovnaní s SCA. SIA z kuracích peria za predpokladu priemernú hodnotu stravy každého jednotlivého Chick je po celú dobu rastu peria v období chick odchovu [18], [19]. Na rozdiel od toho SCA údaje predstavujú sériu "snap-výstrely" (v tejto štúdii každých 5 až 7 dní) pokrmu, že jeden z dvoch rodičov nakŕmiť svoje mláďa [3]. Naša štúdia naznačuje, že SIA tkanív, ktoré integrujú diéty na dlhé časové obdobie sú prirodzene menej variabilné ako SCA danej podobnú veľkosť vzorky a sú vhodnejšie pre posudzovanie medziročné rozdiely v kuracích stravy. Napríklad, dva-source Siar modely použité v našej štúdii boli schopní identifikovať medziročný a druhovo špecifické rozdiely v relatívnej hojnosti rýb a kryl v strave nie sú úplne evidentné pomocou SCA. Pri predbežných informácií na dravých druhovej skladby je obmedzená, ako je mimo obdobia rozmnožovania, na reprezentatívnej zdroj korisť izotopov zmiešavacích modelov môže poskytnúť dôležité informácie o morských vtákov diét, keď nič iného je známe [24]. Avšak, naše výsledky tiež naznačujú, že rozdiely v dravých druhovom zložení v rámci trofickej alebo funkčnými skupinami môže maskovať významné rozdiely v zložení stravy, ktoré by nemohli byť zrejmé z izotopov hodnôt alebo miešanie modelových predpovedí s použitím iba reprezentatívne zdroje korisť. Tento výsledok bol najviac viditeľný pri skúmaní účinku rybieho koristi δ 15N hodnoty na kuracích perie δ hodnotách 15N a dva-source izotopovej zmiešavacie modelov použitých v našej štúdii. Napríklad Chinstrap tučniak perníkový perie δ hodnoty 15N nelíšil rokov (tabuľka 2). Okrem toho 95% dôveryhodnosť intervaly diétneho odhadu z dvoch-source SIAR modelu pomocou p. antarcticum pri parametrizácii naše dva multi-source Siar modely, ktoré sme použili otolitové údaje pre výber vhodných zdrojov rýb korisť zahrnúť do jednotlivých modelových druhov (tabuľka S1 a S2). Avšak, naše počiatočné multi-source SIAR model (Model 3) mal problémy presne odhadovať individuálne druhové zloženie rybej časti tučniakov diét kvôli všeobecnej podobnosti δ 13C a δ hodnoty 15N medzi mnoho z druhy rýb zahrnuté ako korisť zdroja (obr. 3). Antarktickej druhy rýb všeobecne spotrebujú Krill a ďalších druhov rýb a vzhľadom na ich podobnej úrovni tropický tieto druhy rýb majú tendenciu mať podobné δ 15N hodnoty [45].
) a Gentoo ( P. papua
) tučniaky a zvýrazniť spôsoby integrácie dát SCA pre zvýšenie presnosti odhady zloženie stravy s využitím SIA. Odhady SCA biomasy boli vysoko variabilné a podcenil dôležitosť mäkkým telom koristi, ako ryby. Two-source, izotopové zmiešavanie modelové predpovede boli menej variabilné a možno identifikovať medziročný a druhovo špecifické rozdiely v relatívnom množstve rýb a krilu Penguin diéty nie sú úplne evidentné pomocou SCA. V kontraste, multi-source izotopové zmiešavacie modely mali problémy s odhadujúci diétne príspevok druhov rýb okupačných podobných trofické úrovne, bez kultivovanosti použitie SCA odvodených otolitové dáta. Celkovo možno povedať, naša schopnosť sledovať medziročný a druhovo špecifické rozdiely Penguin diétach s využitím SIA bol rozšírený o integráciu dát SCA na izotopovej zmiešavacie režimy tromi spôsobmi: 1) výber vhodných zdrojov korisť, 2) Váhové kombináciou deutériom podobné korisť v dvoch -source miešania modely a 3) rafinácie predpovedal príspevky isotopically podobného koristi v multi-source modely
pre každú trofickú alebo funkčnej skupiny [24], [27], [28]. Ďalej, zatiaľ čo štúdie morských vtákov diét s využitím SIA sa stáva bežným javom, niekoľko štúdií porovnávajúcej súbežné kvantitatívne odhady zloženie stravy medzi SCA a SIA [28], [29]. Okrem toho je tiež spoločné porovnaní dát SIA sa SCA dravých početnosti výskytu namiesto viac príslušných odhadov masovými o zložení stravy, odvodených od SCA [30], [31], [32].
) a Gentoo tučniak viac ako dvoch chovných sezón v Kapskom Shirreff ( P. papua
), Livingston ostrov, Antarktída (62 ° 28'S, 60 ° 46'W) používať oba programu inovácií a SCA. Podobne ako u iných antarktických morských vtákov, Pygoscelis
tučniak diéty sú všeobecne zložené z zooplanktón, predovšetkým antarktického krilu ( Euphausia superba
) a mäkkým telom, vyššie trofické korisť druhy, ako rýb [ ,,,0],33]. Ako chick-správať diéty boli dobre študované pomocou SCA na tomto mieste, že poskytuje vynikajúce prípadovú štúdiu pre porovnanie s SIA [34] [35] [36]. Snažíme sa lepšie pochopiť relatívna výhody oboch metód a poukazujú na využitie SCA informovať izotopové modelov mixážnych lepšie kvantifikovať diéty morských vtákov s využitím SIA
tučniak kurčatá, 2) porovnať prediktívne schopnosť dvoma zdrojmi (krilu vs. ryby) lineárne zmiešavanie modelom medzi tými, s použitím reprezentatívne druhy rýb a tí, ktorí využívajú od a priori
priemerne druhov a rok špecifické hodnoty rýb, a 3) hodnotí metódu a posteriori
integrujúce SCA údaje pre lepšie objasnenie taxonomické zloženie rybie časti diét pomocou multi-source Bayesiánského modelu miešania.
otoliths získaných sme merali náhodný časti vzorky 20-75 P. antarcticum
otoliths na vzorku a tieto hodnoty použité k odhadu rekonštituovanú hmoty tohto druhu.
[44], [45].
stabilný izotop analýzy
štandardné hodnoty boli založené na viedenskej Peedee Belemnite (VPDB) pre ó 13C a atmosférickej N 2 pre ó 15N.
izotopovej miešanie modely
, ktorý sa bežne vyskytujú v Pygoscelis
tučniak diéty ako "rybie" zdroj [6]. Model 2 používa druhov a rok špecifický "ryby" δ 13C a δ 15N hodnoty vypočítané průměrováním delta 13C a ó 15N hodnoty rôznych druhov rýb vážených ich relatívnej percenta z riedeného rybie hmotu (tabuľky S1 a S2).
) a všetky druhy rýb v našej koristi knižnici zistené otoliths v žalúdočného obsahu každý druh "(tabuľka S1 a S2). Model 4 je a posteriori
informovaného modelu, kedy máme obmedzený výsledný zadnej priťahuje k tým, v ktorom bol relatívny význam jednotlivých druhov rýb zaradil v súlade s množstvom jednotlivých druhov určených prostredníctvom otolitových analýzy. Pri modeli 4, máme obmedzený zadné čerpá iba na tie, u ktorých bol odhad pomerná výška príspevku z najrozšírenejších rýb korisť na základe obnoveného rybie hmotnosťou väčšou ako odhadované pomerných príspevkov druhej najhojnejšia ryby korisť, a na druhej najhojnejšia ryby korisť väčšie ako tretia najhojnejšia a tak ďalej pre všetky druhy rýb. U oboch úvodných (Model 3) a informoval (Model 4) multi-source modely sme tiež zhrnuté výsledky naprieč ryby koristi a odhaduje proporcionálne príspevku každého druhu rýb na rybích časti (tj. S vylúčením pancierovky) Penguin stravy. Pri všetkých modeloch SIAR sme začlenené Pygoscelis
tučniak perie δ 15N a δ 13C faktory diskriminácie [47] a bežal 1 milión iterácií, zriediť 15, s počiatočnou vyradenia prvého 40.000 výsledný v 64.000 zadnej kreslí.
výsledky
Izotopové podpisy kuracích peria a korisť
Two-source Siar modelov
izotopové hodnoty ako zástupca "rybí" zdroje (Model 1) a variantu, že použité rok a druhovo špecifické vážené "rybí" izotopové hodnoty (model 2), ako predpovedal, že Tučniak somárov mláďatá spotrebovaný relatívne menej kryl a ďalší ryby ako Chinstrap tučniakov v oboch rokoch (tabuľka 3). Avšak pri skúmaní modelu 95% dôveryhodnosť intervaly tieto dve modelové varianty sa líšili v ich schopnosti detekovať druhovo špecifické, medziročný rozdiely v zložení stravy. Kým obe dve-source SIAR modelových variantov predpovedal, že Gentoo Penguin chick strava obsahovala vyššie percento krilu v roku 2008, Model 2 zistený väčšie množstvo rýb v tučniak čiapočkatý kuracích diét počas roka 2008 v porovnaní s rokom 2009 (tabuľka 3). Two-source Siar modelových variantov predpokladá vyšší príspevok rýb v kuracích diéty oboch druhov tučniakov v porovnaní s odhadmi zloženie diétnych odvodených od SCA vlhkej hmoty (obr. 2). Odhady SCA boli tiež oveľa variabilnejšie než SIAR modelových predpovedí pre Gentoo tučniak mláďa stravy. Okrem toho, SCA odvodené odhady priemerného príspevku krilu a rýb v strave oba druhy poklesol von z našich modelov dvoch zdrojových Siar 95% horné a dolné vierohodnosti intervaly, v danom poradí (obr. 2, tabuľka 1 a 3).
informovaný multi-source Siar modelu (Model 4) fungovala lepšie, než počiatočné viac zdrojov SIAR modelu (Model 3) pri odhadovaní druhové zloženie rybie časti kuracích stravy. Kým Model 4 je predikcia priemernej relatívny pomer jednotlivých druhov rýb v tučniak čiapočkatý kuracích diét mierne líšila od odhadov zo otolitových rozpustené hmoty (× 2 5 = 14.55, p = 0,0125), výsledné intervaly vierohodnosti 95% v porovnaní sa znížili o 53,5 ± 17,2% pre model 3 (rozsah: 33.2-82.2%, tabuľka 4, obr. 3). Okrem toho, Model 4 predikcie zo strednej relatívny pomer jednotlivých druhov rýb v strave Tučniak somárov mláďatá 'bol podobný odhadov otolitových obnoveného hmoty (χ 2 4 = 3,40, p = 0,4949). Navyše, výsledné dôveryhodnosť intervaly boli 95% v porovnaní znížili o 52,0 ± 27,7% na model 3. (Rozsah :. 17.4-76.2%, tabuľka 4, obr 3)
Two-source, Siar modely
ako ryba korisť zdroj (Model 1) prekryté rokoch (tabuľky 3). Naproti tomu 95% dôveryhodnosť intervaly dve-source SIAR modelu s využitím ročnej a druhovo špecifické vážené hodnoty "fish" (Model 2) naznačujú väčšiu hojnosť rýb v tučniak čiapočkatý kuracích diét v roku 2008 v porovnaní s rokom 2009, ktorý bol potvrdený otolitových odvodený, rekonštituovanú priemerná hmotnosť ryby. V roku 2008, ryba časť tučniak čiapočkatý kuracích diét bola zložená zo šiestich druhov rýb s odhadovaným ó hodnote 15N 7,9 ± 0,7 promile, zatiaľ čo P. antarcticum
(δ 15N: 9,4 ± 0,5 ‰) bol jediný druh rýb v roku 2009 diétach (tabuľky S1 a S2). Aj keď to 1,5 ‰ Rozdiel je malý v porovnaní s 4.6-6.1 ‰ rozdiely medzi rybami a kryl, stačilo zmiasť medziročné porovnanie tučniak čiapočkatý kuracích stravy v našej štúdii.
Multiple-source, Siar modely
 Najlepšie 4 veci, ktoré by ste mali vedieť o probiotikách
O probiotikách asi viete. Sú to malé, „priateľské“ chrobáčiky, ktoré podporujú vyvážené a zdravé črevá. Čo možno neviete, je, že ich výhody presahujú zdravie čriev!* Vaše telo je plné dobrých aj zlýc
Najlepšie 4 veci, ktoré by ste mali vedieť o probiotikách
O probiotikách asi viete. Sú to malé, „priateľské“ chrobáčiky, ktoré podporujú vyvážené a zdravé črevá. Čo možno neviete, je, že ich výhody presahujú zdravie čriev!* Vaše telo je plné dobrých aj zlýc
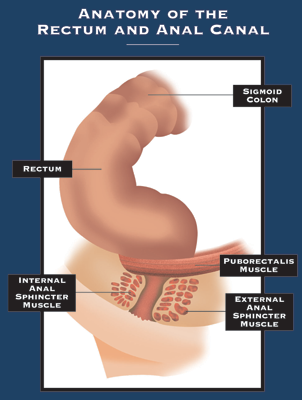 Análne fistuly
Čo je análna fistula? Fistula je abnormálne spojenie medzi dvoma orgánmi alebo dvoma povrchmi tkaniva. Análna fistula (fistula-in-ano) je spojenie medzi análnym kanálom a kožou. Aby sme pochopili prí
Análne fistuly
Čo je análna fistula? Fistula je abnormálne spojenie medzi dvoma orgánmi alebo dvoma povrchmi tkaniva. Análna fistula (fistula-in-ano) je spojenie medzi análnym kanálom a kožou. Aby sme pochopili prí
 Príznaky žalúdočného vredu
Máte pálčivú bolesť v žalúdku sprevádzanú nafukovaním, nedostatkom chuti do jedla a pálením záhy? Ak áno, môžete mať čo do činenia s peptickým vredom. Peptické vredy sú vredy, ktoré vznikajú vo výstel
Príznaky žalúdočného vredu
Máte pálčivú bolesť v žalúdku sprevádzanú nafukovaním, nedostatkom chuti do jedla a pálením záhy? Ak áno, môžete mať čo do činenia s peptickým vredom. Peptické vredy sú vredy, ktoré vznikajú vo výstel