Včasih smo visoko prepustnost pyrosequencing za označevanje z želodcem in vsebine črevesja microbiomes za Crassostrea virginica Navedba. Kralj GM, Judd C , Kuske CR Smith C (2012) Analiza želodec in črevesje Microbiomes vzhodnega Oyster ( Crassostrea virginica Urednik: Josh Neufeld, University of Waterloo, Kanada Prejeto: 8. junij, 2012; Sprejeto: 5. november 2012; Objavljeno: 12. december 2012 To je odprtega dostopa člena se brez vseh avtorskih pravic, in se lahko prosto reproducirati, distribuirati, prenašati, spreminjati, nadgrajevati, ali kako drugače uporablja vsakdo za vse zakonite namene. Delo je na voljo pod Creative Commons CC0 javni domeni predanost Financiranje:. Avtorji priznavajo, NSF dodelitve OCE-1043126 in GoMRI-LSU sredstva za GK za podporo GK, CS in CJ. 454 Titanium zaporedja niso bila zagotovljena z Los Alamos National Laboratory laboratorij Režija raziskave in razvoj Grant (LDRD) do CRK (20080464ER). Med financerji imel nobene vloge pri oblikovanju študije, zbiranje in analizo podatkov, sklep, da se objavi, ali pripravi rokopisa nasprotujočimi si interesi.. Avtorji so izjavili, da ne obstajajo konkurenčni interesi Uvod Eastern ostriga, Crassostrea virginica Mnogi Študije so obravnavale druge vidike ostrig-bakterije interakcij. Cristispira Kljub študij-patogenov, povezanih in prstne odtise zgoraj povzetih in potencialni pomen bakterij za pridobitev ostriga hranil, obstaja presenetljivo malo informacij o ostrig želodca in črevesja microbiome raznolikost. Čeprav so vrednosti pH želodčnih in črevesnih tkiv podobno, ter tranzitnih delcev krat relativno kratko (1-2 h) med aktivno hranjenje [18], ni jasno, ali obstajajo značilne skupnosti v vsebini teh tkiv; je prav tako jasno, kako bi lahko microbiomes spreminja v populaciji ali med prebivalstvom. Za reševanje teh vprašanj, smo dobili dva niza trojnih živali, po en niz od HACKBERRY Bay Lake Caillou v obalnih Louisiani Poleti 2010. Ta dva geografsko ločena mesta (Barataria Bay in Terrebonne Bay, v tem zaporedju) predstavljajo gospodarsko pomemben vir ostrig in izkušnje podobne ureditve slanost in spremenljivost [19]. Smo ločeno zbrali na želodcu in črevesju vsebino in zaporedje PCR pomnožili 16S rRNA gene, ki uporabljajo platformo pyrosequencing (Roche Diagnostics 454 Titanium). Rezultati so pokazali bistveno razlikovanje med želodčnih in črevesnih microbiomes živali iz enem mestu (Lake Caillou), vendar nekoliko manj diferenciacije za drugo spletno stran (HACKBERRY Bay). Predvsem Mollicutes predstavljal > 80% vseh bakterijskih sekvenc v želodcu microbiomes jezera Caillou ostrig, vendar < 10% HACKBERRY Bay ostrig. Želodec Otus tudi Actinobacteria, Chloroflexi, Firmicutes, Planctomycetes, Proteobacteria in Spartobacteria. Chloroflexi, Mollicutes, Planctomycetes in Spartobacteria lahko obsega domnevnega jedro želodec microbiome, medtem ko Chloroflexi, Firmicutes, α-Proteobacteria in Verrucomicrobia bi lahko prispevale k domnevni jedro črevesja microbiome. Primer zbiranje Ostrige so bili zbrani 4. avgusta 2010 od HACKBERRY Bay, majhnem zalivu meji Barataria Bay, Louisiana, ZDA. Ta stran je bila vpliva olja iz razlitja nafte na ploščadi Deepwater Horizon [20]. Treh izvodih ostrige so potekale na ledu (< 6 h) za začetno predelavo v Louisiana morje Grant Oyster valilnico Grand Isle, LA, ZDA. Zunanji ventili temeljito očistiti za odstranjevanje površinske kontaminacije, in nato previdno odprli zapusti žival nedotaknjen. Želodčne vsebine posameznih živali so vzorčnih z uporabo 23-merilnik igle in 1-cm 3 brizg, da dobimo približno 0,2 cm 3 tekočina, ki je bila prenesena v sterilnih 1,5 cm 3 mikrocentrifugirkah. GUT vsebine so bili pridobljeni z lociranjem črevo posameznih živali in nato previdno ekstrudiranje epigastru materiala iz anusa v sterilne 1,5 cm 3 mikrocentrifugirkah. Želodec in črevesje vsebina transportirali na ledu v laboratorij na Louisiana State University (GVŽ), kjer je DNA izpisanih uporabo MoBio Powermax komplet za ekstrakcijo tal (MoBio Laboratories, Inc., Carlsbad, CA), po navodilih proizvajalca z dodatkom zamrznitve (pri -80 ° C, 10 min) /tajanju (pri 60 ° C, 5 min), cikel ponovi trikrat. Drugi sklop ostrig zbranih 1. septembra 2010 od Caillou Bay (Caillou Lake), Louisiana, ZDA so bili obdelani podobno, z izjemo, da so živali, ki se prevaža na ledu na LSU laboratorij pred vzorčenjem želodec in črevesje vsebine. Ta stran je bila vpliva razlitja Deepwater Horizon nafte tudi. Dovoljenja vzorčenja ni bila potrebna za nobeno stran. Analiza DNK DNK izvlečki iz vseh vzorcih so bile s PCR namnožijo z Platinum visoko zvestobo DNA polimeraze (Life Technologies Corp, La Jolla, CA), v 25 ul reakcije, ki uporabljajo standardne protokole z izjemo 68 ° C temperature podaljška, in premazi 515f in 806r modificiranega s črtne kode in adapterji za sekvenciranje uporabljajo pyrosequencing platformo titanovega kemije [21] Roche 454. Vsak reakcijska mešanica je vsebovala 11,5 ul vode, 2,5 ul 10X visoko zvestobo pufra (Life Technologies Corp, La Jolla, CA), 0,75 xl 100 mM dNTP, 1 ul MgSO 4, 5 ul 0,5 mg ml -1 BSA, 1.5 ul za vsako 515f in 806r primerji, 0.2 ul visoko zvestobo DNA polimeraze (Life Technologies Corp, La Jolla, CA) in 1 ul ekstrakta DNK. Reakcijski zmesi smo denaturirali za 3 min pri 94 ° C, čemur sledi 26 ciklov 94 ° C za 1 minuto, 1 min pri 54 ° C in 2 minuti pri 68 ° C, s podaljškom koraku 10 min pri 68 ° C, po cikli so bili popolni. Trojne reakcije za vsak vzorec smo združili in nato končno zmes pripravljena za sekvenciranje z dodajanjem amplikonov iz vsakega vzorca v enakih mas. Pyrosequencing je opravila zaporedja objektu Los Alamos National Laboratory, kar skupno 237,842 surovega bere s povprečno dolžino 295 bp. Zaporedja so bili predloženi v strežnik MG-RAST kot 4501864.3-4501873.3 (http://metagenomics.anl.gov/linkin.cgi?project=1994). Raw zaporedij s kakovostnimi rezultati so bili obdelani s tremi cevovodov. PANGEA [22] je bil uporabljen za primerjavo filogenetske sestavo vzorcev, za katere so Otus razvrščene uporabo Megablast z referenčno zbirko podatkov, ki vsebuje 170,273 celovečernih 16S rRNA zaporedij genov pri bakterijah in Archaea izolira. Raw se glasi so bili pregledani z uporabo privzetih vrednosti (povprečna ocena kakovosti, 20, minimalna dolžina 100 bp) [22]. Bere so umiti na osnovi črtne kode, ki so bili okrašeni pred Megablast. Zaporedja so bili dodeljeni domene /deblo, razred /Da /družino in ravni rodov in vrst, v tem zaporedju, uporabljajo mejne vrednosti podobnost z 0,8, 0,9, 0,95 in 0,99 [22]. Zaporedja niso razvrščene po Megablast so bili zbrani v Otus temelji na istih pragov podobnosti. PANGEA ustvaril tudi drugo analizo, pri kateri so vsi vzorci sestavljalo enako število glasi; ti normirane vzorčne podatkovni nizi so bili izdelani s pomočjo zaporedij naključno izbranih brez nadomestitve iz prvotne pregledanimi vzorčne datoteke. Sestavki želodčnih in črevesnih vzorcev smo primerjali s pomočjo glavnih komponent analize po odpravi enojčkov (sekvence zastopane le enkrat v celotnem CCD), in po odstranitvi cianobakterije in evkariontskih sekvence (kloroplastov in mitohondrijski 16S rRNA iz alg celic v želodce in črevesje). Od preostalih sekvenc, opredeljenih na ravni deblu ali manj reprezentativne zaporedja za Otus obračunavanje ≥0.1% celote so kurator ročno uporabo Megablast v GenBank. Vse sekvence, ki Pangea misidentified prerazporedil po potrebi Plinovod CloVR [23] je bila uporabljena s svojimi privzetimi nastavitvami (npr Povprečna ocena kakovosti, 25, minimalna dolžina 100 bp). Ustvariti analize, ki temelji na taksonomske pripadnosti (tj sestava vzorca) in zaporedje filogenija. V ta namen CloVR uporablja hibridni cevovod je sestavljen iz Mothur sub-rutino, ki se uvrščajo sekvence z bazo podatkov PRP, in QIIME sub-rutine za različne statistične analize. Po odstranitvi cianobakterije in evkariontske sekvence, Otus razvrščena po CloVR ki predstavljalo ≥ 0,1% vseh preostalih glasi bili podvrženi ročno ohranjevanje kot zgoraj. Usklajena reprezentativnih sekvence za tajne, kustosinja Otus so bili nato uporabljeni za analizo Fast UniFrac (http://bmf2.colorado.edu/fastunifrac/), ki temelji na-soseda pridružili drevo kot vhod. Mothur plinovod [ ,,,0],24] je bila uporabljena s strožjim vrednostim kot druge platforme za zaporedje obrezovanje (tj premika okno 50 bt s povprečno oceno kakovosti 35; minimalno dolžino, 100 bp). "Razvrsti" funkcijo cevovoda Mothur je bila uporabljena za identifikacijo sestave vzorca za Otus predstavlja ≥ 0,1% podatkovne zbirke po odstranitvi cianobakterije in evkariontskih sekvence. Preostali sekvence smo kuriral kot je opisano zgoraj. Te kuratorski sekvence plus manjše Otus razen enojčkov so bile uporabljene za ustvarjanje indeksov raznolikosti za vzorce, ki so neodvisne od taksonomske identifikacije (npr Shannon, inverzno Simpson je in indeksi ravnost). pre- obdelavo rutine od zaposlenih v tej študiji treh cevovodov povzročilo izrazito različne zaporedne številke za analizo (tabela S1). PANGEA dala največji prebrati številko (199,592), in Mothur prinesla najmanj (45.626). Zaporedja so najtesneje povezane z cianobakterij in evkariontih (kloroplasti in mitohondrijskih 16S rRNA genov) prevladujejo obrezuje zbirk podatkov (> 70%), ne glede na njihovo velikost (tabela S1). Te sekvence so bili izločeni iz nadaljnjih analiz. Singleton zaporedja predstavljal 0,5% (CloVR) na 5,9% (Pangea) zbirk podatkov po predobdelave; Te sekvence so bile odpravljene tudi za čim manjši vpliv napak zaporedja. Himere zaporedja niso bile ugotovljene pri jo Pangea, vendar se zdi, da predstavljajo le majhen del (< 0,2%). Celotnega zaporedja, (tabela S1), ki temelji na rezultatih iz CloVR in Mothur več vzorcev pojavil dosledno . Relativna številčnost Otus kot odstotek števila zaporedij analiziranih je pokazala, da Lake Caillou oyster želodec in črevesje microbiome skladbe bistveno razlikuje (slika 1a;. Tabele 1, 2). Manjše število Mollicute Otus prevladujejo nekdanji, medtem ko Chloroflexi (večinoma Caldilineae), Firmicutes, γ-Proteobacteria in Verrucomicrobia (Spartobacteria) prevladujejo pozneje. Vsi trije cevovodi pokazala tudi razlike med HACKBERRY Bay ostrig želodca in črevesja microbiomes (slika 1a;. Tabelah 1, 2), vendar razlike so manj izraziti, kot so v Lake Caillou ostrige. Razlike med HACKBERRY Bay želodčnih in črevesnih microbiomes predvsem posledica skromne spremembe v več linij (npr Chloroflexi, Firmicutes, α-Proteobacteria, δ-Proteobacteria, Planctomycetes in Spartobacteria). Poleg tega je vsaka od cevovodov pokazala izrazite razlike med microbiomes obeh populacij iz HACKBERRY Bay in jezera Caillou. Najbolj opazne razlike je prišlo med obema sklopoma želodčne microbiomes, z nekoliko manj razlikovanje med črevesno microbiomes (sl. 1a, Miza 1, 2). Kljub veliko podobnosti, PANGEA, CloVR in Mothur izhod razlikujejo v pomembnih vidikih. Glede na CloVR in Mothur, PANGEA opredeljena manj Proteobacteria, Mollicutes in Verrucomicrobia v HACKBERRY Bay ostriga želodcu microbiomes in manj Actinobacteria, Chloroflexi, Planctomycetes in Verrucomicrobia v črevesni microbiomes. PANGEA tudi dosledno zabeležili večji odstotek "neuvrščene" sekvencah kot storil CloVR ali Mothur; PANGEA ni odkrila 60% HACKBERRY Bay oyster želodčne sekvenc, ki presegajo ravni domene (tabele 1, 2). so opazili tudi razlike v taksonomske pripadnosti v najbolj obilne Otus (tabela 3). PANGEA, CloVR in Mothur vseh prijavljenih Planctomycetes kot eden od dveh enako najbolj bogatih Otus v HACKBERRY Bay ostriga želodčne microbiomes, vendar posebne pripadnosti v Planctomycetes razlikovala. V pripadnost drugega OTU tudi razlikovala, vključno s Firmicute (Pangea), Spartobacteria (CloVR) in Mollicute (Mothur). Poleg tega, PANGEA poročali zaporedje v zvezi z Mycoplasma mobile Analize sestave (debel in razredi) na 284, razvrščenih Otus (sl. 1b) je razkrila vzorce, ki nekoliko oddaljila od tiste, ki temeljijo na relativni številčnosti debel in razredi med vsemi sekvenc (sl. 1a). Najprej so bile razlike med želodčnih in črevesnih microbiomes v mestu in po vsej strani, ki temeljijo na OTU sestavi manj izrazita kot tiste, ki temeljijo na frekvencah pojavljanja (sl. 1a v primerjavi z 1b). To je bilo očitno pri večjih (npr Chloroflexi, Firmicutes, γ-Proteobacteria, δ-Proteobacteria in Planctomyces) in manjše (npr Archaea, β-Proteobacteria in Spartobacteria) prispevajo k OTU sestave (sl. 1b). Po drugi strani je odstotek prispevka nekaj debel in razredih do tajnih Otus močno zastopani glede na njihovo številčnost v podatkovnem nizu zaporedja, medtem ko so drugi debel in razredi bistveno premalo zastopane (sl. 1a, b). Mollicutes so močno zastopani v HACKBERRY Bay in Lake Caillou želodčne microbiomes, vendar premalo zastopane v črevesni microbiomes. Chloroflexi in Planctomyces so zastopani tudi v Lake Caillou ostrig črevesju in HACKBERRY Bay oyster želodčnih in črevesnih microbiomes, medtem ko α- in β-Proteobacteria so premalo zastopane v vseh microbiomes (sl. 1a, b). Podobna vzorec ko se je filogenetski sestava vseh Otus ki se je zgodil v združenih HACKBERRY Bay in Lake Caillou želodca microbiomes v primerjavi s sestavo Otus ki so se zgodile ali so bile v skupni rabi (SHR-s) čez obe strani, so opazili. Zlasti so bili Chloroflexi, Mollicutes, Planctomyces in Spartobacteria zastopani med SHR-S Otus (sl. 2). Podobno je primerjava Otus, ki se pojavljajo v združenih HACKBERRY Bay in Lake Caillou črevesja microbiomes z deljenim črevesju v Otus (SHR-G), je pokazala, da so bile Chloroflexi, Firmicutes, α-Proteobacteria, Planctomyces in Verrucomicrobia zastopani (sl. 2). Število SHR-S Otus (44) je precej manjše od števila SHR-G Otus (112), pri čemer slednji predstavljali skoraj 40% vseh razvrščenih Otus in še večji odstotek tistih najdemo v črevesju microbiomes (Tabela 4). Otus ki nedvomno prišlo v želodcu ali črevesju microbiomes tako HACKBERRY Bay in ostrig populacije Lake Caillou (SHRU-s, SHRU-G) predstavljen nek drug podskupina. SHRU-S microbiome je predstavljal le 5 od 44 SHR-S Otus v samo 3 debel /razrede, in so predstavljali le 2,1% od 284 skupno Otus opredeljena v kolektivni želodca in črevesja microbiomes (tabela 4). Nasprotno pa sta bila SHRU-G microbiome s 44 112 SHR-G Otus zastopana v 12 debel /razrede, in je predstavljal 15,5% vseh opredeljenih Otus (preglednica 4). Sestava SHR-S in SHRU-S microbiome Otus razlikuje bistveno, medtem ko so razlike med microbiomes SHR-G in SHRU-G omejena na manj debel in razredih (sl. 2). Poleg variabilnosti med želodca in črevesja filogenetske sestave so microbiomes spreminjala med ponovitvene ostrig iz vsake strani. Za nekatere debel in razrede, je bila relativna številčnost podobni med ponovitvami, in spremenljivost (izražena kot standardna napaka srednje vrednosti) je bila podobna za vsako od treh cevovodov (glej na primer Mollicutes in alfa-Proteobacteria pri želodčnih in črevesnih microbiomes, v tem zaporedju; tabela 1, 2). Vendar pa v mnogih primerih ponovitev bistveno spreminjala, in obseg variabilnosti razlikovala med cevovodov. Mollicutes v črevesju microbiome HACKBERRY Bay, na primer, je bilo v samo 1 od 3 ponovitev po CloVR in Mothur opazili, in so bili nesorazmerno bogat v eni ponovitvi po Pangea (tabela 2). variabilnost med ponovitvami je bil ujet z analizo skupin (sl. 3) in analize glavnih komponent (PCA) rezultatov CloVR uporabo UniFrac razdalje (sl. 4a, b), in tudi PCA relativnih številčnosti zaupnih Otus (sl. S1). Izsledki analize grozdov z uporabo tehtanega UniFrac meritev je pokazala, da je jezero Caillou želodec razmnožuje in HACKBERRY Bay gut ponavlja vsak oblikujejo različne grozde, in da so bile posamezne ponovitve relativno blizu v daljavo. Preostali želodec in črevesje vzorci so bili veliko manj usklajena, s ponovitvami rešiti, na večje razdalje. Netehtano UniFrac PCA je pokazala, da Lake Caillou gut microbiomes združeni na osi ena in dva, vendar vzporednih so bili drugi microbiomes veliko bolj razpršena, čeprav so razlike med lokacij in med črevesja in želodca ostala očitna (sl. 4a). Tehtano UniFrac PCA, ki šteje Relativne prisotnosti Otus, je pokazala, da Lake Caillou želodec in HACKBERRY Bay gut ponavlja vsak oblikovana razmeroma ozkih skupin na obeh oseh, medtem ko so ponovitve za druge microbiomes razpršena (sl. 4b). Dva želodcu microbiomes ostala tudi loči na PCA osi 1, vendar so gut microbiomes združeni (sl. 4b). Mi predstavljamo prve podrobne analize Crassostrea virginica Delne 16S rRNA genska zaporedja, ki izhajajo iz velikega pretoka, pyrosequencing kot je uporabljen v tej študiji kažejo razlike v oyster microbiome sestavi na več ravneh, čeprav nekatere podrobnosti o sestavi razlikujejo s plinovodom je bil izbran za sekvenčno analizo (npr tabele 1 , 2). Oglejte si dodatne informacije S1 za dodatno obravnavo teh razlik, ki ne vplivajo na vzorce razlike med želodčnih in črevesnih microbiomes ali razlike med lokacijami. Na splošno so rezultati kažejo velike razlike med želodčnih in črevesnih microbiomes, in med želodec microbiomes živali iz HACKBERRY Bay Lake Caillou (npr slika 1a, b in 2;. Tabele 1, 2). Poleg tega microbiome sestavki posameznih ponovitvenih živali razlikujejo (sl. 3, 4). Razlike med želodčnih in črevesnih microbiomes verjetno odraža podrobnosti prebavnega sistema, vendar razlike med lokacij in med ponovitvami kažejo, da bi microbiome sestava odziv na lokalne dejavnike, in morda genetskih razlik med posamezniki. Analogno variabilnost so poročali pri drugih živali [28], [29]. Na podlagi pogostosti OTU pojavljanja, želodec microbiome ostrig iz Louisiane lahko obstajajo ob najmanj dveh državah. Mollicutes najtesneje povezane s Mycoplasma fiziološki in ekološki pomen teh oyster želodčne microbiomes je negotova. Prevlada, ki Mollicutes ali Planctomycetes je nekoliko nenavadno glede na druge microbiomes [28], [30] - [32], čeprav Mollicutes pojavi bogat v prebavnem žlezi Sydney Rock Oyster je ( Saccostrea glomerata Doslej genomske dokazi ponuja nekaj spoznanj, saj je genetsko repertoarja Mycoplasma mobile Vloga Planctomyces v prebavni sistem je prav tako negotova. Čeprav so ekološko pomembna člani morskega bacterioplankton, funkcionalno raznovrstnih in povezane z alg, nevretenčarjev in vretenčarjev [48], [49], se navadno pojavijo pri relativno nizki številčnosti v črevesni microbiomes (< približno 5%) [30] - [32]. Vendar pa rezultati te študije kažejo, da je neznani pogoji HACKBERRY Bay oyster želodec korist Planctomycete orožja za (tabela 1). To je skušnjava, da špekulirajo tukaj, saj imajo drugi drugje [50], ki Pirellula Oyster Gut Microbiome oyster gut microbiome pristaniščih bolj speciose ali OTU bogati skupnost kot pa želodec microbiome na osnovi opazovanih vrst (s obs) in ACE in Chao1 ocenjevalcev raznolikosti (tabela 5). Ti indeksi prav tako kažejo, da želodci in čreva iz Lake Caillou ostrige pristanišče manj Otus kot HACKBERRY Bay ostrig. Tako OTU bogastvo giblje med ostrig tkiv (na primer, želodčnih in črevesnih), kot je bila dobro dokumentirana za človeško microbiome [53], vendar se zdi, da tudi razlikuje med populacijami. Vir razlik v bogastvu med ostrig populacije ni znan. Razlike v bogastvu Ne glede na črevo microbiome ni nujno bolj raznolika kot v želodcu microbiome temelji na Shannon in inverzna indekse Simpson in ravnost cenilko, od katerih vsaka so podobni za črevesja microbiome Lake Caillou in dveh HACKBERRY Bay microbiomes (tabela 5). Ta podobnost kaže, da je v nekaterih primerih struktura ostrig microbiome raznolikosti (bogastva in ravnost) neodvisna od prebavnega sistema in phylotype sestavo. Nasprotno, vsi indeksi raznolikosti za želodčne microbiome Lake Caillou so bistveno nižje kot pri HACKBERRY Bay želodcev, in nižja od obeh črevesno microbiomes kot dobro. To lahko pripišemo prevlado v Lake Caillou ostrig želodcev v Mollicute Otus (npr Tabela 5 in sl. 1a). Sestava črevesno microbiomes iz Louisiana ostrig razlikuje od drugih školjk, in iz druge morske in ne-morskih živali (sl. 5). Gruenthal [54] je pokazala, da Proteobacteria prevladujejo (> 80%) so gut microbiomes v Kaliforniji črne barve ( Haliotis cracherodii Te razlike v sestavi med črevesno sistemov izhajajo iz učinkov več v stiku spremenljivk, vključno črevesja arhitekture, prebavne fiziologije, prehrana, in v kolikšni meri gosti in microbiomes razvilo složno [61 ] - [63].
, Velikonočni ostrige, dobljena iz dveh mest, eno v Barataria Bay (HACKBERRY Bay) in drugi v Terrebonne Bay (Lake Caillou), Louisiana, ZDA. Želodec microbiomes v ostrige iz HACKBERRY Bay so večinoma prevladujejo Mollicutes so najtesneje povezana z Mycoplasma
; bolj bogati skupnost s Planctomyctes prevladujejo prišlo v Lake Caillou ostrig želodcev. Gut skupnosti za ostrige iz obeh strani, razlikuje od želodca skupnosti, in gojila relativno raznolik skupek phylotypes. Phylotypes najtesneje povezana s Shewanella
in Chloroflexi seva prevladoval v Lake Caillou in HACKBERRY Bay žive mikroorganizme v črevesju oz. Medtem ko mnogi člani želodca in črevesja microbiomes je zdelo, da je prehodni ali oportunisti, je bil domnevni jedro microbiome določene na podlagi phylotypes ki so se zgodili samo v vseh želodcu ali črevesju vzorcev. Domnevna Jedro želodec microbiome sestavlja 5 Otus v 3 debel, medtem ko je domnevni jedro gut microbiome vsebuje 44 Otus v 12 debel. Ti rezultati skupaj razkrila nove mikrobne skupnosti v sistemu ostriga prebavnega, funkcije ostrigarja microbiome so večinoma neznani. Primerjava microbiomes iz Louisiana ostrig s bakterijske združbe so poročali pri drugih morskih nevretenčarjev in rib je pokazala, da so bili mehkužcev microbiomes bolj podobni med seboj, kot da microbiomes v Polychaeten, decapods in rib
) iz obalnih Louisiana, ZDA. PLoS ONE 7 (12): e51475. doi: 10,1371 /journal.pone.0051475
, ki je znan po svoji tržne vrednosti in pomena kot "ekosistemski inženir" [1] - [3]. Obseg je bilo napisanih o njegovi biologiji in ekologiji, vključno interakcij z bakterijami in drugimi mikrobi. Velik del te literature je poudaril bolezni [4], [5] in prisotnost človeških patogenov, še posebej Vibrio parahaemolyticus
in V
. vulnificus
[6] - [9].
je bila opredeljena kot simbiotskimi povezano z kristalni nakupu, mehkužcev prebavne strukturo [10]. Stappia
(zdaj Labrenzia
) je bil izoliran pri C
. gigas
in C
. virginica
in slednji vpleten kot antagonist za etiološko sredstvo mladoletniškega Oyster bolezni [11]. Študije kultura odvisna od sta značilna Vibrio
in drugih rodov povezana z razsutem stanju živali in določenih tkiv [6] - [8], [12] [13], vključno z določitvijo "avtohtonih" bakterije v C . gigas
hemolimfa [14], [15]. Takšne študije so tudi pokazale, da je vzhodno razlitje Sredozemlje olje ni vplival ostriga povezano bakterije [16]. Kultura neodvisen študije so dokumentirani vzorcev raznolikosti med različnimi populacijami in tkiv, v primerjavi-drstenja postavljeno in divje živali, in ugotovila, da je ε-Proteobacterium, Arcobacter
, kot pomembno prispeva k mikrobne združbe čilskega ostriga Tiostrea chiliensis
[17].
Materiali in metode
Zaporedje Analiza
Rezultati
kot najbolj obilne OTU za HACKBERRY Bay ostrig črevesno microbiomes, medtem ko so preostali cevovodi poročali sev Chloroflexi (tabela 3). V nasprotju s tem, trije cevovodi so pokazali veliko bližje soglasje za Lake Caillou vzorcev: vse ugotovi, da OTU tesno povezana z M
. mobile
je bil najpogostejši v želodcu microbiomes, in OTU tesno povezan s Shewanella
sp. je bil najpogostejši v črevesni microbiomes. Dva Shewanella
izolatov poročajo, Mola 59 (PANGEA) in THt8-1 (CloVR in Mothur), so bile enake kot položajih nukleotidnih analiziranih. Vendar pa je Shewanella
sp. THt8-1 in Shewanella
sp. Mola 59 smo izolirali iz kopenskih rastlinskih in morskih virov oz.
Pogovor
želodec in črevesje microbiome skladbe. Velikost vzorca (trojne živali za vsako od dveh mestih) in z enim meja ekstrapolacija čas vzorčenja rezultatov, ampak zagotavljajo številna nova spoznanja. Prejšnje študije so poudarili obdelovalnih člani črevesja skupnosti, cele živali, patogeni (človeških in ostrig), ali posebne skupine, ki lahko prispevajo k prebavo, na primer, Cristispira
[5], [8] - [11] [13] [25]. pristopi gojenja brez so razkrili Arcobacter
(ε-Proteobacteria) kot pomembno prispeva k mikrobnih združb celih čilskih ostrig, Tiostrea chilensis
, vendar niso poročali celotna tkiva posebna združenja [17 ]. Hernádez-Zarate in Olmos-Soto [26] so uporabili skupinsko specifičnih FISH in PCR za ugotavljanje bakterij v C
. gigas
tkiva, vendar pa ni zaporedje PCR amplikonov ali poročila relativno obilje posameznih filogenetskih skupin. Pred kratkim je PCR in DGGE študija C
. je virginica
poročali o prostorskih in sezonskih razlik celih microbiomes živalskih dveh populacij iz Maina (ZDA), vendar filogenetsko sestava ni bila ocenjena kakovostno ali količinsko [27], niti niso razlike med posameznimi živalmi je opisana.
Oyster Želodec Microbiome
pretežno prevladujejo male sekvence (> 80%), države, ki jo Lake Caillou ostrige zastopane (slika 1a;. Mize 1, 3). Noben drug razred prispeva več kot okoli 2%. Planctomycetes prevladujejo (23% -33%) namestnik izjava, da od HACKBERRY Bay ostrig (slika 1a;. Tabele 1, 3) - vendar več drugih skupin se pojavljajo tudi v želodcih teh ostrig na skromni številčnosti, na primer, Chloroflexi (8 %), Firmicutes (9% -11), Mollicutes (5% -9%), Proteobacteria (5% -12%) in Verrucomicrobia (3% -14%). Poleg tega, sta prav tako bogat Otus, ki pripadajo različnim debel (Planctomyces in bodisi Firmicutes, Tenericutes ali Verrucomicrobia) prevladujejo HACKBERRY Bay ostrig želodci na ravni vrste (evolucijska razdalja = 0.03; tabela 3). Delež zaupnih Otus odpade na različne debel in razredih je tudi v skladu z dveh različnih držav, za želodcu microbiome (sl. 2), čeprav so razlike manj izrazite, za to meritev kot pri ocenah, ki temeljijo na frekvenčnih sestave. UniFrac PCA (tehtano in netehtano) in cluster analize zagotavljajo dodatno podporo za "dve državni" koncept (sl. 3, 4).
) in v črevo na morsko uho, Haliotis discus hannai
[33], [34]. Mollicutes so poročali tudi ostrig gut Kelih celic, ki temelji na mikroskopskem dokazov [25], in dokumentiran za druge vretenčarje in ribjega drobovja, ki temelji na kulturi in molekularne ekološke metode [30], [35] - [42]. Sicer relativno malo znanega o njihovih zvez z nevretenčarjev sistemov prebavo. Dejansko ekološke vloge Mollicutes bolj na splošno še vedno negotove, z nekaterimi poročili patogenezo v izbranih rib in nevretenčarjev [43] -. [46], vendar druga poročila, ki kažejo neko obliko komenzalizmu [30], [39]
je takson najtesneje povezana z ostrig Otus, je omejena v svojem obsegu [47]. M
. mobile
congenerics v ostrig želodcev lahko preprosto razmnoževali s pomočjo substrata, ki ga gostitelja ali drugih mikrobov med prebavo; podobni predlogi so bili na računu Mollicute združenja s hladno vodo koral [39]. Kljub temu pa je možnost, da bi Mollicutes složno prispevajo k njihovi gostitelji ne more zavrniti.
-podobne člani ostrig microbiome izkoriščajo Sulfatiran alg polisaharide za rast, saj so opazili številni geni domnevno kodirajo sulfohydrolase encimov v Rhodopirellula Baltica
genoma [51], in ker bi sulfatirani polisaharidi biti splošno zaužije ostrig kot posledico potrošnje fitoplanktona. Možnost uporabe sulfatirane polisaharide bi tako pojasni razloge za Planctomycete izobilju. Na žalost, filogenetski odnosi med R
. Baltica
in planctomycete Otus opredeljene v tej študiji ne zadoščajo za podporo takšne sklepe. Kljub temu pa vse Blastopirellula
, Pirellula
in Rhodopirellula
izolatov značilna do danes uporabljajo široko paleto preprostih brez žveplovih sladkorjev [48], [49], [ ,,,0],52], vsaj nekatere, ki se lahko pojavijo v oyster prebavnem traktu, kot je biomasa alg hidroliziran.
) in belo morsko uho ( H
sorenseni
. ); Actinobacteria, Chloroflexi, Planctomyces in Verrucomicrobia zdi, da je odsoten od obeh. Huang et al. [41] kažejo, da Mollicutes in δ-Proteobacteria prevladujejo črevo malega morsko uho. Cardoso sod. [55] Poročilo, ki Bacteroidetes in Firmicutes prevladujejo črevesje za polže, polž, ahatnik
. Firmicutes skupaj z Bacteroidetes, Proteobacteria in Actinobacteria prevladujejo poguma drugih nevretenčarjev (npr termitov hranjenje tal [56] in ščurki [57]) in vretenčarji (npr rastlinojede morske ribe [58], amur, [31], [ ,,,0],59] in primati [60]), medtem ko Mollicutes prevladujejo poguma nekaterih rib [30] [36]. V nasprotju s tem, Proteobacteria predstavljajo le okoli 20% sestave črevesne od ostrig v tej študiji, Chloroflexi, Planctomyces in Verrucomicrobia vsak relativno bogata in Actinobacteria, Bacteroidetes, Firmicutes in Mollicutes vsak prispevati približno 10% ali manj (sl. 1a;. Tabela 3)
 Kako se lahko hitro znebim divertikulitisa?
Kako se lahko hitro znebim divertikulitisa?
 Puščajoče črevesje in mikrobna disbioza bi lahko prispevali k nevihti citokinov pri hudo bolnih primerih COVID-19
Puščajoče črevesje in mikrobna disbioza bi lahko prispevali k nevihti citokinov pri hudo bolnih primerih COVID-19
 Študija povezuje porabo fermentirane zelenjave z nizko smrtnostjo zaradi COVID-19
Študija povezuje porabo fermentirane zelenjave z nizko smrtnostjo zaradi COVID-19
 Peptid pajkovega strupa bi lahko pomagal ustaviti bolečine pri sindromu razdražljivega črevesja
Peptid pajkovega strupa bi lahko pomagal ustaviti bolečine pri sindromu razdražljivega črevesja
 Največji mit o spremembah in kaj storiti glede tega
Največji mit o spremembah in kaj storiti glede tega
 Zakaj ponovno testiram za SIBO
Zakaj ponovno testiram za SIBO
 Lazy Man's Moules Mariniéres (SCD/Paleo Friendly)
Kaj? Sem len človek. Spoštujem francosko kuhinjo in vse ostalo. Pripravijo prekleto dobro hrano. Toda to ni bila romantična večerja, bil je naporen torkov večer. Torej, moral sem narediti tisto, kar n
Lazy Man's Moules Mariniéres (SCD/Paleo Friendly)
Kaj? Sem len človek. Spoštujem francosko kuhinjo in vse ostalo. Pripravijo prekleto dobro hrano. Toda to ni bila romantična večerja, bil je naporen torkov večer. Torej, moral sem narediti tisto, kar n
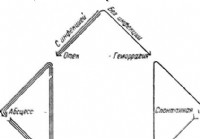 Razvrstitev akutnega pankreatitisa - Diagnoza akutnega abdomena
Ponujenih je veliko klasifikacij akutnega pankreatitisa. Najpogosteje so klasifikacije temeljile na naravi procesa in morfoloških spremembah trebušne slinavke. Tako je bila apopleksija trebušne slinav
Razvrstitev akutnega pankreatitisa - Diagnoza akutnega abdomena
Ponujenih je veliko klasifikacij akutnega pankreatitisa. Najpogosteje so klasifikacije temeljile na naravi procesa in morfoloških spremembah trebušne slinavke. Tako je bila apopleksija trebušne slinav
 Je carski rez dober za zdravje otrok?
Nedavna študija, ki jo je objavil Inštitut za ekonomske raziskave VATT, kaže, da je nenačrtovan CS opravil za indikacijo, ki bi se ji morda lahko izognili, ima vzročno-posledično povezavo z astmo že o
Je carski rez dober za zdravje otrok?
Nedavna študija, ki jo je objavil Inštitut za ekonomske raziskave VATT, kaže, da je nenačrtovan CS opravil za indikacijo, ki bi se ji morda lahko izognili, ima vzročno-posledično povezavo z astmo že o