kurkumín zvyšuje expresiu SCF /c-kit cez tlmiace oxidačný stres a aktiváciu NF-kB v žalúdočných tkanivách diabetických gastroparézy krýs
abstraktné
pozadia
Diabetes mellitus je spojená s mnohými druhmi komplikácií. Nedávne štúdie ukázali, že oxidačný stres a zápalové reakcie majú kľúčovú úlohu v patogenéze diabetickej gastroparézy. Kurkumín je známe, že majú antioxidačné a protizápalové vlastnosti. V tejto štúdii sme skúmali vplyv kurkumínu na diabetickej žalúdočnej motilitu v modeli Sprague Dawley krýs o diabetes mellitus 1. typu.
Metódy
Samce SD potkany boli rozdelené do kontrolnej skupiny, kontrolná skupina, ktoré kurkumín, diabetik skupinu a diabetická skupina, ktoré kurkumín. Diabetes sa indukuje intraperitoneálnou injekciou streptozotocínom. Kurkumín (150 mg /kg) bol uvedený do žalúdka počas 6 týždňov, a bola meraná hladina glukózy v krvi a telesnej hmotnosti. Žalúdky boli vyňaté pre analýzu žalúdočného vyprázdňovanie sadzieb a úrovní oxidatívneho stresu. NF-kB, I-kB, a faktor kmeňových buniek (VVP) /hladiny c-kit proteín boli hodnotené analýzou Western blot, zatiaľ čo apoptóza intersticiálnych buniek Cajal (ICC) bola hodnotená farbením TUNEL.
Výsledky
kurkumín liečených diabetických krýs vykazovali výrazne zlepšené rýchlosti vyprázdňovania žalúdka [(59,4 ± 7,5)%] v porovnaní s diabetických krýs [(44,3 ± 5,7)%], ako aj znížené hladiny MDA [21,4 ± 1,8 (nmol /mg) vs
27,9 ± 2,1 (nmol /mg)], a zvýšená aktivita SOD [126,2 ± 8,8 (jednotiek /mg) vs.
107,9 ± 7,5 (jednotiek /mg)]. Na druhej strane, žalúdočné vyprázdňovanie hladina v kontrolnej skupine nebolo významne odlišné od kontrolnej skupiny, ktorí boli liečení kurkumínový. Okrem toho, kurkumín liečených diabetických krýs vykazovali významne zvýšené hladiny SCF /c-kit proteínu v žalúdočnej tkanive, inhibuje degradáciu I-kB a aktiváciu NF-kB, a zníženie indexu ICC apoptózy [(26,2 ± 4,1)% vs
(47,5 ± 6,2)%], v porovnaní s diabetickou skupinou.
Záver
ošetrenie Kurkumín zlepšiť vyprázdňovanie žalúdka tým, že blokuje produkciu oxidačného stresu, zrušenie NF-kB prenos signálu a zvýšenie expresie SCF /c-kit v krysy s diabetickou gastroparesu.
Kľúčové
kurkumín Diabetická gastroparézy Oxidačný stres NF-kB faktora kmeňových buniek /c-kit intersticiálna bunky Cajal z úvodu
diabetickej gastroparesu je ochorenie zažívacieho traktu, definované ako oneskorené vyprázdňovanie solídne jedlo, a je vidieť v 30-50% pacientov s typu 1 alebo diabetes mellitus 2. typu [1]. Pacienti často prítomné hornej gastrointestinálne príznaky, ako je skoré sýtosti, chudnutie, nadúvanie, bolesti brucha, nevoľnosť, vracanie sa vyskytujú často, a zhoršenie kontrolu glykémie, a choroba vážne ovplyvňuje kvalitu života pacientov.
Gastroparézy sa čím ďalej častejšie považovaná za významnú zdravotný problém. Hoci gastroparézy ovplyvňuje mnoho diabetikov na celom svete, sú nielen možnosti liečby veľmi obmedzené, ale aj liečby, ktoré sú k dispozícii pre diabetickej gastropatia sú často neúčinné. Patrí medzi ne lekársku terapiu, žalúdočné elektrickú stimuláciu, chirurgické terapiu a nutričné podporu. Prípravky, ktoré majú gastroparesu patrí metoklopramid, domperidón, cisaprid a erytromycín. Pričom tieto látky boli použité pre liečbu gastroparézy, boli hlásené ako len obmedzenú účinnosť, a mnoho pacientov nemôže tolerovať ich, pretože ich vedľajšie účinky. U diabetických pacientov s refraktérnym gastroparesu, vysokofrekvenčné žalúdočné elektrická stimulácia pomocou trvalo implantovaného systému výrazne zlepšili symptómy hornej časti tráviaceho traktu a predstavuje alternatívne chirurgická liečba pre pacientov. Chirurgické zákroky, ako je gastrektómii a antrectomy sú posledné možností pre liečbu, byť kontroverzné a potrebujú ďalšie štúdie [2]. U pacientov s ťažkým gastroparézou, ale normálne malé čriev, jejunostómie plniaca trubica môže byť použitá. To poskytuje nutričné podporu, sa nelieči gastroparézy [3]. Diabetická gastroparézy je všeobecne najlepšie liečené, a to len u pacientov s ťažkou diabetickou gastroparézou, ktorí neuspeli v reakcii na medikamentóznu liečbu by mal podstúpiť ďalšie terapiou. Tak, zistenie lieky, ktoré vykazujú účinnosť v liečbe diabetickej gastroparézy je nutné.
Diabetická gastroparézy je spojená so stratou intersticiálnych buniek Cajal (ICC), ako u ľudí tak modelových zvierat [4]. ICC možno identifikovať v tkanive štítkoch s antiséra proti kinázy súpravy na receptor tyrozín, proteín exprimovaný na ICC. Kit a jeho kmeňových ligand bunkového faktor (VVP) sú tiež dôležité faktory pre prežitie ICC [5]. Zvýšený oxidačný stres spojený s diabetom môže viesť k strate alebo poškodení ICC u myší, zatiaľ čo oxidačný stres vedie k poškodeniu a strate ICC sietí a rozvoj oneskoreného vyprázdňovania žalúdka [6]. Výsledky diabetes v zvýšeného oxidačného stresu a hrá dôležitú úlohu v patogenéze diabetických komplikácií [7]. Nadprodukcia reaktívnych foriem kyslíka (ROS) vedie k oxidačnému poškodeniu, vrátane peroxidácie lipidov, oxidácie proteínov a poškodenie DNA, čo môže viesť k bunkovej smrti. Okrem toho, ROS je známe, že pôsobí ako druhí poslovia k aktivácii transkripčných faktorov, ako je nukleárna faktor kappa B (NF-kB). ROS vznikajúce pri namáhaní v žalúdočnej tkanive môže spustiť aktiváciu NF-kB signálne kaskády; skutočne, vyprázdňovanie žalúdka u diabetických potkanov je spojená s aktiváciou NF-kB sprostredkovanej zápalom.
kurkumín (1,7-bis (4-hydroxy-3-metoxyfenyl) -1, 6-heptadien-3,5- dión), je hlavná účinná zložka kurkumy izolovať z rastliny kurkumy longa L. kurkumín je multifunkčný molekula s významnými regulačné účinky na rakovinu [8], zápal [9] a diabetické súvisiace ochorenia, ako je diabetická retinopatia [10], diabetická nefropatia [11], a diabetes kognitívne dysfunkcie [12]. Kurkumín je silný lapač reaktívnych foriem kyslíka a dusíka, ako sú hydroxylové radikály a radikály oxidu dusičitého [13]. Niekoľko štúdií preukázalo, že kurkumín inhibuje aktiváciu NF-kB prozápalové signálne dráhy [14]. Medzitým, NF-kB transkripčné faktory regulujú rad dôležitých fyziologických procesov, vrátane zápalu a imunitných odpovedí, bunkového rastu a prežitie, a tak inhibícia NF-kB signalizácia predstavuje životaschopnú stratégiu pre liečbu ochorení [15]. Avšak, tam bol veľmi malý výskum potenciálu kurkumínu na liečenie diabetickej gastroparézy, alebo na jeho farmakologického mechanizmu.
Cieľom tejto štúdie bolo zistiť, či, v Sprague Dawley (SD) potkaním modelu diabetu typu 1 mellitus, stredné dávky [16] kurkumín mohol ochrániť ICC u diabetických krýs s oneskoreným vyprázdňovaním žalúdka znížením oxidačného stresu a inhibovať aktiváciu NF-kB, čo zvyšuje SCF /výraz Kit a normalizáciu oneskorenie vyprázdňovania žalúdka.
materiály a metódy
Experimentálne potkany a indukcia diabetu
Male SD krýs (vek 8 týždňov) boli chované v centre pokusných zvierat, Zhejiang čínskej medicíny univerzity, (Hangzhou, Čína), kde špecificky prostého patogénov (SPF) -Level laboratórium bola schválená Zhejiang provinčné vlády. Všetky potkany boli uložené za podmienok riadenou vlhkosťou (50-60%). Boli udržiavané pod kontrolovanou svetlom (12 h denný /nočný cyklus) s voľným prístupom k vode a hlodavce. Všetky pokusy na zvieratách boli vykonané v súlad s licenciou z provincie Zhejiang Science and Technology Office (Hangzhou, Čína) a so súhlasom etickej komisie živočíšne Zhejiang čínskej lekárskej univerzity. Všetky pokusy zodpovedalo pokyny pre etické správanie pri používaní a ošetrovaní zvierat. Všetko úsilie bolo vykonané, aby sa minimalizovalo stresu k zvieratám.
Krysy na lačno po dobu 12 hodín prešli jednou intraperitoneálna injekcia streptozotocínom (STZ), 50 mg /kg, čerstvo rozpusteného v 100 mM citrátovom sodnom pri pH 4,5. Vek-uzavreté normálne potkany dostávali iba citrátu. Vývoj diabetu bol potvrdený nalačno hladinu glukózy v krvi (FBG) pomocou reagenčnej súpravy (Roche, Shanghai, Čína). Krysy s hladinou FBG vyššou ako 11,1 mm (200 mg /dl), pri 72 hodinách po injekcii STZ boli považované za diabetické krysy. Akonáhle krysy sa stal diabetik, ich hladina glukózy boli merané denne. Experimenty sa začala 8 týždňov po vzniku diabetu umožniť gastroparézy rozvíjať.
Experimentálneho dizajnu
Osem týždňov po vzniku diabetu, zvieratá bola rozdelená do štyroch skupín, tj normálne ovládanie samcov laboratórnych potkanov (skupina I), diabetická samca potkany (skupina II), kurkumín doplnené diabetickej samce potkanov (skupina III), a kurkumín doplnené riadiacou samce potkanov (skupina IV). Krysy v skupinách III a IV (n = 10 každý) boli ošetrené denne curcumin (150 mg /kg, i.g.) po dobu 6 týždňov. Kurkumín (čistota väčšia ako 95%) z Fusong kraj prírodnej Biotechnology Company Ltd. (Jilin, Čína), bol suspendovaný v 0,5% w /riešenie v sodné karboxymetylcelulózy (CMC-Na). Krysy v skupinách I a II (n = 10 každý) dostala iba 0,5% roztoku CMC-Na. Po liečbe po dobu 6 týždňov, boli zvieratá usmrtené v etyl-éteru anestézii, bola odobratá krv od stehennej žily krvácania a sérum sa oddelí. Žalúdky boli rýchlo odstránené a vzorky tkaniva odobraté od zvierat, boli skladované pri -80 ° C až do spracovania pre biochemické testy. Vyprázdňovanie žalúdka, oxidatívny stres, aktivácia NF-kB, ICC a SCF /výraz Kit v žalúdkoch boli tiež študoval na konci šiesteho týždňa liečby (14
th týždeň cukrovky).
Vyprázdňovanie žalúdka štúdie
bola použitá štandardná metóda na prípravu fenolovej červene markeru jedlo, ako bolo opísané skôr [17]. Vyprázdňovania žalúdka sa stanoví za použitia modifikácie minulosti hlásené postupom [18]. Krysy bol ponechaný voľný prístup k vode až do 3 hodín pred podaním sondou kurkumínu. Roztok 0,1% (w /v) fenolu vo vodnej sodné soli karboxymetylcelulózy (1,5% w /v) bol použitý ako testovací jedla. Kurkumín bola daná 1 h pred orálnym podaním testované jedla. Dvadsať minút po podaní testovanej jedlá, boli krysy usmrtené. Žalúdky boli potom vystavené pomocou laparotomie a odstránená. Krysy ošetrené vehikulom (fyziologický roztok, 0,9% roztok chloridu sodného, 0,2 ml) boli usmrtené okamžite po orálnom podaní testovanej jedla a fenolovej červene obsahu žalúdka bol považovaný za štandard (100%). Vyňaté žalúdky sa nareže v 40 ml roztoku hydroxidu sodného (0,1 N) a obsah sa rozpustí. A 1-ml podiel z pracích roztokov sa pridá ku 2 ml kyseliny trichlóroctovej (7,5% w /v) k vyzrážaniu proteínov. Po odstredenia (2500 x g
) počas 20 minút, pridá sa 1 ml supernatantu sa 1 ml roztoku hydroxidu sodného (1 N) vyvinúť maximálnu intenzitu farby. Absorbancia pri 558 nm roztoku sa merala pomocou spektrofotometra (U-1080, Hitachi Ltd., Tokyo, Japonsko). Vyprázdňovanie žalúdka bola vypočítaná podľa tohto vzorca: vyprázdňovanie žalúdka %
=
1
-
X
/
Y
×
100
,
kde X je absorbancie fenolovej červene, zostávajúce v žalúdku 20 minút po podaní fenolovej červene a Y je priemerná absorbancie fenolovej červene dostal sa z žalúdkov kontrolných myší bezprostredne po fenolovej červene podaní.
Stanovenie MDA a hladiny SOD v hladkých svaloch žalúdočnej dutine
žalúdky boli vyrezané, zvážené a bezprostredne potom zmrazil pri -70 ° C. Zmrazené tkanivá od každého potkana sa homogenizuje v ľadovo chladnom pufri fosforečnanu (KCl 140 mM, fosfát 20 mM, pH 7,4) a centrifugován pri 3000 x g počas 10 minút
. Obsah malonaldehyde (MDA), index pre peroxidácie lipidov, sa stanoví za použitia metódy kyselinou thiobarbiturovou (TBA) s miernou modifikáciou [19]. Stručne, 100 ul tkanivového homogenátu bolo zmiešané s 200 ul pracovného roztoku, ktorý obsahuje 0,37% TBA. Zmes sa inkubuje pri teplote 100 ° C počas 15 minút a potom sa ochladí. Extrahovať MDA, pridané 375 ul N-butanolu, nasledovalo vortexování po dobu 10 sekúnd. Po odstredení, horná N-butanol vrstva sa prenesie do sklenenej skúmavky. Absorbancia fáza butanolu bola meraná pri 532 nm. obsah MDA je vyjadrená ako nmol /mg proteínu. Pre meranie aktivity SOD, 20 ul tkanivového homogenátu bolo zmiešané s 200 ul reakčnej zmesi obsahujúce nitroblue tetrazoliumchlorid (NBT, 750 um), a inkubované po dobu 20 minút pri teplote 37 ° C. Absorbancia pri 560 nm bola meraná. Enzýmová aktivita je vyjadrená ako jednotky /g proteínu a 1 jednotka enzýmu je definovaná ako množstvo enzýmu potrebné na inhibíciu redukcie NBT o 50% meraní.
Semikvantitatívny RT-PCR z SCF a c-kit
proximálny žalúdka (40-50 mg) bola zozbieraná a sliznice sa odstráni preparáciou. Tkanivo sa mechanicky homogenizuje v RNase-free podmienok a disociované Tripura izolácie činidla (Roche, Švajčiarsko) na ľade po dobu 5 minút a bola extrahovaná celková RNA podľa návodu výrobcu. Celková RNA bola kvantifikovaná spektrofotometricky pri 260 a 280 nm, pričom pomer A260 /A280 v rozmedzí od 1,8 do 2,0. Integrita RNA bola overená elektroforézou na agarosovém gélu, nasledované farbením etidiumbromidom a RNA bola použitá pre okamžité reverzné transkripciu alebo skladované pri -80 ° C vo vode bez RNázy. Dvojstupňový reverznej transkriptázy-PCR (RT-PCR) bola vykonaná s použitím ACE-α- syntézu prvého vlákna cDNA kit Revertra (Tiangen Biotech, Beijing, Čína) a 2 × Taq PCR master mix (Tiangen Biotech), v súlade s pokyny výrobcu. Pevne stanovená RNA (0,5 ug) bol reverznej transkripcii. Glyceraldehyd-3-fosfát dehydrogenázy (GAPDH) bol vybraný ako vnútorný štandard. Všetky PCR priméry boli navrhnuté a syntetizované Biosune (Shanghai, Čína). Tieto sekvencie primérov a dĺžky PCR produktov sú nasledovné: GAPDH, vpred: 5 '- GACAACTITG-GCATCGTGGA-3 ', reverzné: 5 '- ATGCAGGGATGATGT-TCTGG-3 ' , 150 bp; SCF, vpred: 5 '- GGA CTT CAT GGT GGC ATC TG-3 ', reverzné: 5 '- GCC CTT GTA AGA CTT GAC TG-3 ', 285 bp; a c-kit, vpred: 5 '- GTG GTT AAA GGA AAC GCT CG-3 ', reverzné: 5 '- CAT ACA TTT CAG CAG GTG CG-3 ', 400 bp. Reakčná podmienky boli optimalizované pre každý z génov zmenou žíhacie teplotu (58 ° C počas GAPDH, 58 ° C počas SCF, a 62 ° C pre c-kit) a rôznymi počty cyklov PCR [20]. Produkty PCR boli rozdelené na 2% agarosovém gélu pri 100 V po dobu 30 minút. Gélové obrazy boli zobrazované na displej z tekutých kryštálov monitora ultrafialového presvietením systému PhotoDoc-ito vybaveným CCD kamerou (Bio-Rad Gel Doc 2000, Bio-Rad Laboratories, Hercules, CA) a zachovaná pre analýzu intenzity. Relatívna hladiny mRNA vybraných génov boli vypočítané ako pomer k GAPDH prejavu.
Úrovne SCF, c-Kit, IkappaB a NF-kB hodnotená pomocou analýzy western blot
asi 50 mg tkaniva boli prevzaté z v blízkosti proximálnej žalúdok. Tkanivá boli narezané na malé kúsky asi 0,25 cm 3, homogenizuje, a rozpustné vo radio-Imunoprecipitácia test lyzačním pufra (Hushang Biotechnology, Shanghai, Čína), obsahujúcim 50 mM Tris, 150 mM NaCl, 1% NP-40, 0,5% deoxycholát sodný, 1% dodecylsulfát sodnom, 1 mM PMSF, orthovanadičnan sodný, fluorid sodný, kyselina etyléndiamíntetraoctová, a leupeptinu pri teplote 4 ° C počas 30 minút. Homogenát boli centrifugovány pri 12 000 x g
dobu 5 minút pri teplote 4 ° C a supernatanty boli zbierané a použité ako celkovej bielkoviny. Rovnaká množstvo proteínu sa podrobí elektroforéze na SDS-PAGE na 10% alebo 15% polyakrylamidových géloch. Proteíny boli prenesené na nitrocelulózové membrány (Hushang Biotechnology) o polosuchou blotting. Membrány boli blokované s Tris-pufrovanom fyziologickom roztoku 0,1% Tween 20 (TBST) s obsahom 5% mlieka po dobu 60 minút pri teplote miestnosti, a potom sa immuno-blotting s vhodne zriedeným primárnymi protilátkami pri 4 ° C cez noc. Po premytí trikrát TBST, boli bloty inkubovali s HRP-konjugovanou sekundárnou protilátkou po dobu 60 minút pri teplote miestnosti. Potom sa komplexy boli vizualizované za použitia zobrazovacieho systému iChemi XR (WD9431A, Bomeike Biotechnology, Čína), s chemiluminiscencie činidiel (Tigsun Biological Science Technology, Tianjin, Čína). Semi-kvantifikácia bola vykonaná pomocou Množstvo Jeden softvér, verzia 4.6.2 (Bio-Rad). Boli použité nasledujúce protilátky: anti-SCF, 1: 200, c-kit, 1: 200, anti-IkappaB a anti-NF-kB P65 a GAPDH., 1: 500 (Hushang Biotechnology, Shanghai, Čína)
kvantifikácia NF-kB -DNA väzba elektroforetické mobility shift testu
jadrové extrakty boli pripravené z žalúdka lyzátu, ako bolo opísané skôr [21]. Aktivácia NF-kB bola stanovená pomocou elektroforetického posunu mobility testu (EMSA), a nukleárnej translokácii NF-kB bola hodnotená ako je popísané skôr (GS009, Beyotime Institute of Biotechnology, Shanghai, Čína) [22]. Pre NF-kB Väzbové reakcie, 5 ug extraktu jadrového proteínu sa inkubuje pri teplote miestnosti po dobu 20 minút s reakčného pufra obsahujúceho 20 mM HEPES, pH 7,9, 50 mM KCI, 0,1 mM EDTA, 1 mM DTT, 5% glycerol, 200 ug /ml BSA a 2 ug poly (dl-dC). Potom, 32P-značený dvojvláknové oligonukleotid (1 ng ≥1 x 105 cpm), ktorý obsahuje NF-kB viažuci konvenčné sekvencie (5 '- GGCAACCTGGGGACTCTCCCTTT-3 ") bol pridaný k reakčnej zmesi po dobu ďalších 10 minút pri teplote miestnosti. Reakčné produkty boli frakčnom na nedenaturujícím 6% polyakrylamidovom gélu po dobu 60 minút pri 350 V, ktorý sa potom suší a podrobí autorádiografiou pri -70 ° C cez noc. Za kompetitívnych testoch, prebytok oligonukleotid (100-násobný molárny nadbytok) súťažiaci bola vopred inkubovaná s jadrovými extraktmi po dobu 20 minút pri teplote miestnosti. Mutant NF-kB oligonukleotid (5 '- GGCAACTGCTCACTCTCCCTTT-3 ") bol použitý pre testu súťaženie. Signály boli analyzované denzitometrické.
Dvojstranná immunofluorohistochemistry s terminálny deoxynukleotidyltransferáza sprostredkovaná dUTP-biotín nick end označovanie testu (TUNEL) a c-kit farbenie
fluorescenčné imunohistochémia pre c-kit (pomocou protilátky z Santa Cruz) a TUNEL farbenie boli vykonávané na 10-um častiach za použitia modifikácie postupu, zverejneného výrobcom súpravy TUNEL (TAC STDT, Trevigen Inc., Gaithersburg, MD). TUNEL-pozitívne bunky boli značené fluoresceínom (Ex 488, Em 505-550), c-kit-pozitívne bunky boli označené Cy3 (Ex 543, EM 560 - 615) a jadrá boli označené DAPI (Ex 364, EM 385 -470). Tkanivové vzorky boli zakotvené v fluorescencie montovacího médiá a vyšetrovaný konfokálna laserovou skenovacej mikroskopie [23]. Približne 200 Bunky boli spočítané na poli, päť polí boli skúmané na snímku a päť rezy boli skúmané v jednej skupine. Percentuálne podiely TUNEL- a c-kit-pozitívny apoptotické bunky boli označené ako apoptotické index (AI) (%). Štatistická analýza
Všetky dáta sú uvádzané ako priemer ± SD. Pre porovnanie dát medzi skupinami zvierat, jednocestné analýzy variance (jednosmerná ANOVA) a porovnávanie Duncan boli zamestnaní. Všetky štatistické testy boli vykonané s použitím SPSS pre Windows, verzia 13.0 (SPSS Inc., Chicago, IL). Rozdiely boli považované za štatisticky významné pri P Hotel < 0.05.
Výsledky
Vplyv kurkumínu na telesnej hmotnosti a krvnej glukózy diabetických krýs
plazmatické hladiny glukózy boli veľmi zvýšená u diabetických krýs (skupina II) [(322 ± 41) mg /dl] v porovnaní s tými, u normálnych kontrolných krýs (skupina I), [(98 ± 12) mg /dl]. Došlo k výraznému Pokles telesnej hmotnosti zo skupiny krýs II [(298 ± 16) g] v porovnaní s krysami skupiny I vekovo [(449 ± 21) g]. Chronické kurkumín liečených diabetických krýs (skupina III) nepreukázali žiadne významné zlepšenie telesnej hmotnosti a žiadne zníženie hladiny glukózy v plazme v porovnaní s krysami skupiny II [skupiny III, telesnej hmotnosti (324 ± 19) g a glukózy (302 ± 38) mg /dL] (tabuľka 1) .Table 1 v krvi hladina glukózy a telesnej hmotnosti do štyroch skupín
Skupiny
skupina aj
skupina II
skupina III
Skupina IV
glukózy v krvi (mg /dl)
98 ± 12
322 ± 41 *
302 ± 38 * #
100 ± 11
Telesná hmotnosť ( g)
449 ± 21
298 ± 16 *
324 ± 19 *
476 ± 27
skupiny I = normálna kontrolné krysy; Skupina II = diabetickej kontrolnej krysy; Skupina III = diabetická + kurkumín liečených potkanov (150 mg /kg); Skupina IV = normálny + kurkumín liečených potkanov (150 mg /kg). Výsledky sú uvedené ako priemer ± SD. * P Hotel < 0,05 (v porovnaní so skupinou I), #P Hotel &0,05 (v porovnaní so skupinou II)
Vplyv kurkumínu na vyprázdňovanie žalúdka
vyprázdňovanie žalúdka výrazne predĺžila na SD potkanov, ktoré diabetes po dobu 8 týždňov. , Množstvo vyprázdňovanie žalúdka v skupine II bol 44,3 ± 5,7%, čo je podstatne nižšia ako v skupine I [(76,2 ± 4,3)%, P Hotel < 0,01], a diabetické krysy prechádza kurkumín liečba [(skupina III),; (59,4 ± 7,5)%, P Hotel < 0,05]. Avšak, žiadny významný rozdiel bol zaznamenaný medzi skupinou I a krýs skupiny IV [(73,3 ± 5,2%), P Hotel > 0,05] (Obrázok 1). Obrázok 1 Vplyv kurkumínu na vyprázdňovanie žalúdka u diabetických krýs. Skupina I = normálny kontrolný krysy (n = 10); : Skupina II = diabetickej kontrolnej krysy (n = 10); : Skupina III = diabetická + kurkumín liečených krýs (n = 10); : Skupina IV = normálny + kurkumín liečených krýs (n = 10). Všetky hodnoty sú vyjadrené ako priemer ± SD. * P
< 0,01 (v porovnaní so skupinou I), # p Hotel &0,05 (v porovnaní so skupinou II), AP Hotel >. 0,05 (v porovnaní so skupinou I)
Kurkumín liečba znížila tvorba MDA, zvyšuje aktivitu SOD
sme skúmali oxidačný stres za použitia niekoľkých metód, pretože glukóza-indukovanej oxidatívny stres je nastolila, že je kľúčovým mechanizmom v chronických diabetických komplikácií. Diabetickej krysy vykazovali zvýšenú hladinu MDA [skupiny I 15,7 ± 1,7 (nmol /mg) vs
skupiny II 27,9 ± 2,1 (nmol /mg), P
= 0,006] a zníženej hladiny SOD [skupiny I 146,2 ± 9.3 ( u /mg) vs.
skupiny II 107,9 ± 7,5 (u /mg), P
= 0,008], je molekulárna marker oxidačného stresu (Obrázok 2). Šesť týždňov liečby znížil kurkumín stupeň MDA up reguláciu [skupiny III, 21,4 ± 1,8 (nmol /mg), P
= 0,033 oproti skupine I a P
= 0,004 oproti skupine II], a zlepšila SOD dole -regulation [Group III 126,2 ± 8,8 (jednotiek /mg), P
= 0,036 oproti skupine i a P
= 0,001 oproti skupine II]. Kurkumín výrazne zvýšenú aktivitu SOD a zníženej hladiny MDA v žalúdku Homogenát. Okrem toho, žiadny významný rozdiel medzi skupinami boli zaznamenané I a skupina IV zvierat (P Hotel &0,05; Obrázok 2). Obrázok 2 Vplyv kurkumínu na MDA a SOD u diabetických krýs. Hladiny MDA bola znížená po spracuje kurkumínu, hladiny SOD sa zvýšila potom, čo spracuje s kurkumín. : Skupina I = normálna kontrolnej skupiny krýs (n = 10); : Skupina II = diabetickej kontrolnej krysy (n = 10), skupina III = diabetická + kurkumín liečených krýs (n = 10); : Skupina IV = normálny + kurkumín liečených krýs (n = 10). Všetky hodnoty sú vyjadrené ako priemer ± SD * p
. ≪ 0,01 (v porovnaní so skupinou I), # p Hotel < 0,01 (v porovnaní so skupinou II), AP Hotel &0,05 (v porovnaní s skupina i).
Kurkumín zvyšuje SCF a hladiny c-kit proteín
reprezentatívne zobrazenie analýzy RT-PCR z SCF v žalúdku tkanivách laboratórnych potkanov v každej skupine sú znázornené na Figure3A. V porovnaní s skupiny I, SCF bol v skupine II výrazne nižšia [(0,44 ± 0,02) vs
(0,69 ± 0,03), P
= 0 0,002], a porovnanie hodnôt v skupine III ukázala, že hladina SCF v kurkumín potkanov bol väčší, ako v skupine II [(0,58 ± 0,04 vs)
(0,44 ± 0,03) P
= 0 .003]. Avšak neboli zaznamenané žiadne rozdiely medzi skupinami I a IV [(0,69 ± 0,03) vs
(0,66 ± 0,03) P
= 0,56] (Figure3B). Western blot analýza (Figure4A, B), sa zistilo, že úroveň SCF v žalúdočnej tkanive od potkanov skupiny II bola významne nižšia ako u potkanov skupiny I a III [pomery (0,33 ± 0,02) vs
(0,47 ± 0,03) a (0,41 ± 0,02), v danom poradí, P Hotel < 0 0,05)]. Avšak, žiadny významný rozdiel bol zistený medzi skupiny IV a skupiny I krýs [(0,45 ± 0,03) vs
(0,47 ± 0,03), P Hotel > 0.05)], čo naznačuje, že kurkumínový zásah mohol účinne obnoviť miestnej úrovne SCF bielkovín v žalúdku tkanivách diabetických krýs. Obrázok 3 Zmeny v SCF /GAPDH a C-kit /GAPDH RT-PCR analýzu. (A) Reprezentatívne obrazy gélovej elektroforézy GAPDH, SCF, a c-kit na krysách v každej skupine. hladiny (B) Skupina relatívnej expresie mRNA SCF a c-kit, s výrobkami kvantifikovať pomer GAPDH. : Skupina I = normálna kontrolnej skupiny krýs (n = 10); : Skupina II = diabetickej kontrolnej krysy (n = 10); : Skupina III = diabetická + kurkumín liečených krýs (n = 10); : Skupina IV = normálny + kurkumín liečených krýs (n = 10). Všetky hodnoty sú vyjadrené ako priemer ± SD. * P
< 0,01 (v porovnaní so skupinou I), # p Hotel &0,05 (v porovnaní so skupinou II), AP Hotel >. 0,05 (v porovnaní so skupinou I)
Obrázok 4 zmeny v SCF /GAPDH a C-kit /GAPDH pomocou Western blot analýzy. (A) Zástupca proteínový pás obrazy SCF, c-kit a glyceraldehydes-3-fosfátdehydrogenázy (GAPDH). hladiny (B) Skupina relatívnej SCF a c-kit proteínu, vyjadrená pomerom k GAPDH, resp. : Skupina I = normálna kontrolnej skupiny krýs (n = 10); : Skupina II = diabetickej kontrolnej krysy (n = 10); : Skupina III = diabetická + kurkumín liečených krýs (n = 10); : Skupina IV = normálny + kurkumín liečených krýs (n = 10). Všetky hodnoty sú vyjadrené ako priemer ± SD. * P Hotel < 0,01 (v porovnaní so skupinou I), # p Hotel < 0,01 (oproti skupine II), AP Hotel >. 0,05 (oproti skupine I)
RT-PCR analýza ukázala, že hladiny expresie pre c-kit v žalúdočnej tkanive boli 0,78 ± 0,04, 0,37 ± 0,02 a 0,59 ± 0,03 pri skupine aj, II a III, v danom poradí. Hladiny c-kit expresie v skupinách II a III potkany boli nižšie ako v skupine I u potkanov. Avšak porovnanie skupín II a III bolo zistené, že hladina c-kit v kurkumín potkanov bol väčší, než u neošetrených diabetických krýs (P
menšie ako 0,05, Figure3B). Podobné výsledky boli demonštrované pomocou Western blot analýzy (Figure4B). Hladiny c-kit proteínu (pomer 0,45 ± 0,03, 0,21 ± 0,02 a 0,34 ± 0,02, respektíve pri skupine I, II a III) bolo v skupine III oveľa vyššia ako v skupine II (P Hotel < 0 0,05, Figure4B), a hladina v skupine III bola nižšia ako v skupinách i a IV. Obe analýzy ukázali, že 6 týždňov curcumin intervencie by mohli významne obnoviť pokles hladín c-kit u diabetických krýs.
Kurkumín inhibuje degradáciu I-kB a aktiváciu NF-kB
správy naznačujú, že zvýšená a chronická oxidatívny stres môže aktivovať alebo rušiť NF-kB aktivity [24]. Skúmali sme, či kurkumín môže viesť k aktivácii NF-kB signalizačnej dráhe. K tomu, sme merali hladiny NF-kB v prítomnosti alebo neprítomnosti kurkumínu na určenie, či sa NF-kB aktivovaný. Po prvé, sme hodnotili hladiny proteínovej I-kB, ku ktorému inaktivovaná NF-kB sa viaže. Zistili sme, že úroveň I-kB v skupine I potkanov bola vyššia ako u potkanov skupiny II, zatiaľ čo hladina u potkanov Skupina III bola nižšia ako u potkanov skupiny II (p 0,01), ale vyššie ako hladiny v skupiny i a IV potkany (p menšie ako 0,05). Potom sme skúmali, či kurkumín môže blokovať NF-kB. Zistili sme, že diabetes (skupina II), viedlo k zvýšeniu hladiny NF-kB [skupiny I, 75,5 ± 7,7 (pg /ml) proti
skupiny II 102,8 ± 12,4 (pg /ml), P = 0,001]
, zatiaľ čo liečba kurkumín [skupiny III 91,3 ± 8,9 (pg /ml)] zabránené tento účinok. Na základe tohto zistenia sme navrhli, že kurkumín potláča aktiváciu NF-kB [skupiny IV 70,9 ± 8,1 (pg /ml), P = 0,026
] (Figure5). Skúmali sme, či kurkumín môže blokovať NF-kB translokáciu do jadra, pretože nukleárna translokácia je rozpoznaný ako bunkové reakcie na stimuláciu ROS a zdá sa, že koreluje s NF-kB sprostredkovanej transkripčný aktivácie. Za týmto účelom posúdenia, sme merali hladiny NF-kB v jadre EMSA. Vývoj diabetu indukovaného zvýšenia hladiny NF-kB v jadre, zatiaľ čo liečba kurkumín zabránené tento účinok (Figure6). Preto sme navrhli, že kurkumín nielen nepriamo potláča aktiváciu NF-kB-viažuci DNA, ale že tiež potláča aktiváciu proti smeru I-kB. Obrázok 5 Vplyv kurkumínu na I-kB a NF-kB. (A) stĺpcový graf bol jasný odraz úrovne I-kB a NF-kB u štyroch skupín. (B) úroveň I-kB /GAPDH bola znížená v skupine II, ale zvýšil o kurkumín. I: Skupina I = normálna kontrolnej skupiny krýs (n = 10); II: Skupina II = diabetickej kontrolnej krysy (n = 10); III: Skupina III = diabetická + kurkumín liečených krýs (n = 10); IV: Skupina IV = normálny + kurkumín liečených krýs (n = 10). Všetky hodnoty sú vyjadrené ako priemer ± SD * p
. ≪ 0,01 (v porovnaní so skupinou I), # p Hotel < 0,01 (v porovnaní so skupinou II), AP Hotel &0,05 (v porovnaní s Všetci autori čítať a schválená konečná rukopis.
 Príčiny bolesti na pravej strane hrudníka
O ľavostrannej bolesti na hrudníku a jej vzťahu k srdcovým chorobám sa veľa hovorí. Ale čo pravostranná bolesť na hrudníku? Hoci sa môže zdať bezpečné predpokladať, že nesúvisí so srdcovým problémom –
Príčiny bolesti na pravej strane hrudníka
O ľavostrannej bolesti na hrudníku a jej vzťahu k srdcovým chorobám sa veľa hovorí. Ale čo pravostranná bolesť na hrudníku? Hoci sa môže zdať bezpečné predpokladať, že nesúvisí so srdcovým problémom –
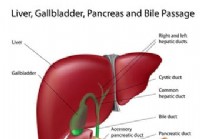 Ochorenie pečene
Diagram priechodu pečene, pankreasu a žlče. Symptómy a príznaky ochorení pečene zahŕňajú slabosť a únavu, chudnutie, nevoľnosť, vracanie a žlté sfarbenie kože (žltačka). Aké fakty by som mal vedieť o
Ochorenie pečene
Diagram priechodu pečene, pankreasu a žlče. Symptómy a príznaky ochorení pečene zahŕňajú slabosť a únavu, chudnutie, nevoľnosť, vracanie a žlté sfarbenie kože (žltačka). Aké fakty by som mal vedieť o
 Aké sú príznaky dumpingového syndrómu? Skoré vs. Neskoré
Symptómy dumpingového syndrómu zahŕňajú nadúvanie, nevoľnosť, vracanie, kŕče v bruchu, slabosť, potenie, závraty, zrýchlený tep, a trasením Dumpingový syndróm, nazývaný aj rýchle vyprázdňovanie žalúd
Aké sú príznaky dumpingového syndrómu? Skoré vs. Neskoré
Symptómy dumpingového syndrómu zahŕňajú nadúvanie, nevoľnosť, vracanie, kŕče v bruchu, slabosť, potenie, závraty, zrýchlený tep, a trasením Dumpingový syndróm, nazývaný aj rýchle vyprázdňovanie žalúd